|
7. DIAGNÓSTICO DO MEIO BIÓTICO
De forma a atender o preconizado pelo Termo de Referência Nº 017/99, o diagnóstico do meio biótico foi dividido em dois itens: ambiente terrestre e ambiente aquático, abordando as características da Flora e Fauna em cada um destes.
Os estudos foram realizados por especialistas da Universidade Federal do Rio de Janeiro - UFRJ, de forma a caracterizar as Áreas de Influência Direta (AID-5km e AID-15km) com base em levantamentos primários e representar a Área de Influência Indireta (AII) através de dados secundários.
7.1. Flora Terrestre- topo
7.1.1. Cobertura Vegetal - topo
Angra dos Reis (23º15’S, 44o12’W) está localizada a 155 km da cidade do Rio de Janeiro e, juntamente com os municípios vizinhos, formam um complexo geográfico e turístico denominado Costa Verde. Possui um litoral bastante recortado e diversos ecossistemas em sua extensão: florestas, manguezais, restingas e costões rochosos.
O clima da região é quente e úmido sem estação seca. A temperatura média anual é de 22,5ºC, sendo a máxima média 25,7ºC em fevereiro e a mínima média 19,6ºC em julho (Bittencourt, 1997). Angra dos Reis apresenta uma nebulosidade elevada (setembro a março); a pluviosidade anual é de aproximadamente 2.240 mm, com maior concentração no verão (Domingues et al.,1976). Os ventos predominantes, durante todo o ano são os do quadrante leste, com maior incidência do sudoeste durante os meses mais frios, de junho a setembro (GEA - Feema).
Devido à topografia, que permite um escoamento rápido das águas para o oceano, os rios da região são de pequeno curso, dos quais o único de vazão considerável é o rio Mambucaba. Na faixa litorânea da região, predominam os solos hidromórficos (Xavier et al., 1988). As planícies arenosas são estreitas, sendo as formadas pelos rios Jurumirim ou da Guarda, Bracuí e Mambucaba, na divisa com o município de Parati, as formações geológicas mais amplas (Domingues et al., 1976).
A cobertura vegetal existente nas áreas de influência do empreendimento, conforme levantamento realizado por meio de fotos aéreas, imagens de satélite e dados secundários, pode ser vista nos mapas de uso e ocupação do solo das áreas de influência, nos anexos do Volume 4 (Diagnóstico Ambiental - Meio Socioeconômico) deste EIA.
7.1.2. Mata Atlântica- topo
Considerando todos os ecossistemas que constituem a biosfera, as florestas tropicais detêm sem dúvida a maior diversidade específica tanto animal como vegetal, sem considerar ainda a contínua descoberta de espécies novas nos mais diversos grupos de seres vivos (Lino, 1992; Mittermeier et al. 1999; Wilson, 1997).
No Brasil, os dois principais biomas de floresta tropical são a Floresta Amazônica e a Floresta Atlântica (Ferri,1980; Rizzini, 1997). O primeiro ocupa a região norte do país, enquanto que o segundo percorre ao longo a costa do Oceano Atlântico, originalmente do Estado do Rio Grande do Sul até o Estado do Rio Grande do Norte (Tabela 1).
Tabela 1 - Valores absolutos e relativos das áreas originais e remanescentes da Mata Atlântica no Brasil (Brasil, 1998).
Clique aqui para visualizar a tabela 1
Atualmente o conceito de Mata Atlântica é abrangente e leva em conta não só a floresta propriamente estabelecida em solos consolidados, dominada por indivíduos de porte arbóreo, mas engloba uma série de ecossistemas associados como os campos de altitude, brejos, pântanos, lagunas, restingas, dunas, praias, rios, estuários, enseadas, sacos, gamboas, ilhas, lajes, coroas, costões e pontões rochosos, manguezais e restingas, com todas as suas variações, ocorrentes juntamente ao longo de nossa costa. A mesma abordagem foi feita pelo Decreto 750 de 10/02/1993 - DOU de 11/02/1993. Sendo assim, adota-se aqui a denominação “Mata Atlântica” para se referir ao ecossistema maior, que envolve os demais citados acima, e a denominação “Floresta Atlântica” para se referir ao ecossistema florestal, outrora tipicamente encontrado em nossas baixadas e encostas litorâneas.
A costa brasileira, originalmente, apresentava-se quase que totalmente ocupada por florestas intertropicais. Atualmente tais florestas foram em sua grande maioria removidas, iniciando-se com o extrativismo direto (o ciclo extrativista do pau-brasil - Caesalpinia echinata Lam.), típico da fase pré colonial, passando pelos ciclos de cultivo de cana de açúcar e café associados à extração de madeiras, com posterior implantação de áreas agropastoris. Segundo Borém & Ramos (2001) todas estas modificações ocorreram sem que houvesse uma preocupação com a proteção ambiental. No entanto, tal afirmação é contestada pela obra de Pádua (2002) que afirma que estas preocupações já existiam pelo menos à partir de 1786, mas na realidade somente em 1937 se criou o Parque Nacional de Itatiaia, e na região em 1971, o Parque Nacional da Bocaina e o Parque Estadual da Serra do Mar em 1977.
Estes ecossistemas têm sofrido assim, fortes pressões antrópicas de modo que hoje se verifica a extinção maciça de espécies e o desaparecimento de comunidades e ecossistemas, que ocasionam profundas modificações na paisagem em escala geográfica.
No Estado do Rio de Janeiro em especial, grande parte da superfície total da Mata Atlântica vem sofrendo diversas formas de alteração, principalmente em decorrência da atividade antrópica. Todavia, é crescente a conscientização da necessidade de conservação e de recuperação ambiental para a própria sobrevivência humana, sendo o estado do Rio de Janeiro, responsável por 39 unidades de conservação, na esfera Federal e Estadual, representando aproximadamente 5.700 km2 protegidos, no domínio da Mata Atlântica.
Esta formação pode ser assim dividida em três grandes núcleos florestais característicos: um que domina no sul do país, com um clima mais ameno e solos mais profundos, onde se destaca no estrato superior a Araucaria angustifolia, o pinheiro brasileiro, mas que também pode ser encontrado em áreas limitadas sobre as Serras do Mar e Mantiqueira, dentro dos Estados de São Paulo, Rio de Janeiro, onde predomina principalmente a Floresta Atlântica de Encosta. O terceiro núcleo está representado pelas florestas que predominam do norte do Espírito Santo até o sul da Bahia, e localizam-se sobre regiões de Tabuleiros Terciários (Ferri, 1980; Garay et al., 1995; Jesus, 1987; Lino, 1992).
A Floresta Atlântica desenvolveu-se sobre a cadeia montanhosa que acompanha quase toda a costa brasileira, avançando para o interior do país em diferentes extensões, variando sua proximidade ou distanciamento à costa. Entretanto o núcleo que recobre os Tabuleiros Terciários apresenta afinidades tanto com a Floresta Amazônica como com a Floresta de Encosta (Peixoto & Gentry, 1990). Com efeito, a estrutura que a Floresta Atlântica de Tabuleiros apresenta é similar à da Floresta de Terra Firme, seja pelo porte das espécies arbóreas, seja pela abundância restrita de epífitas, pteridófitas, etc (Ferri, 1980; Peixoto & Gentry, 1990). Enquanto que a relação observada com a Floresta Atlântica de Encosta se dá por afinidade taxonômica, sendo que ambas sofrem diretamente influência do mar.
No Brasil, a Mata Atlântica já representou cerca de 15% de sua cobertura original, sendo que atualmente encontra-se em sua maior parte substituída por sistemas agropastoris ou florestas secundárias degradadas (Jesus, 1987; SOS Mata Atlântica et al., 1998), sendo que nos últimos cinco anos segundo ISA (2002), chegou-se a uma cifra de destruição de 11% da Mata Atlântica, provocando uma queda de 8,3% para 7,3% dos remanescentes em todos os estágios de Mata Atlântica. A taxa de desmatamento é de três vezes a da Floresta Amazônica, correspondente a um campo de futebol a cada quatro minutos. As matas remanescentes que persistem, encontram-se, em geral, em áreas onde o relevo dificulta o acesso e exploração ou em Unidades de Conservação sejam particulares ou federais. A mesma situação é observada também para seus ecossistemas associados.
Na Tabela 2 a seguir, pode-se observar o ritmo da degradação das formações florestais da Floresta Atlântica no estado do Rio de Janeiro, que levaram, em 500 anos, à redução para cerca de 17% de sua superfície original.
Tabela 2 - Evolução histórica da perda de superfície florestal no estado do Rio de Janeiro ao longo dos anos (SOS Mata Atlântica & INPE, 2001).
| Ano |
Área (ha) |
% cobertura florestal natural em relação à área do estado |
| 15001 |
4.294.000 |
97,00 |
| 19122 |
3.585.700 |
81,00 |
| 19601 |
1.106.700 |
25,00 |
| 19783 |
973.900 |
22,00 |
| 19854/6 |
914.691 |
20,83 |
| 19904/6 |
884.112 |
20,13 |
| 19955/6 |
738.402 |
16,82 |
| 20006 |
734.629 |
16,73 |
1 Fundação Instituto Estadual Florestal do Rio de Janeiro.
2 Campos, Gonzaga de ‑ Mappa Florestal.
3 Magnanini, Alceu; Nehab, Maria Alice & Moreira, José Carlos ‑ 1º Simpósio Brasileiro de Sensoriamento Remoto.
4 Atlas dos Remanescentes Florestais e Ecossistemas Associados do Domínio da Mata Atlântica no período 1985 ‑ 1990. Fundação SOS Mata Atlântica/INPE, 1993.
5 Atlas da evolução dos Remanescentes Florestais e ecossistemas associados da Mata Atlântica no período 1990 ‑ 1995. Fundação SOS Mata Atlântica/INPE/ISA, 1998.
6 Atlas dos Remanescentes Florestais da Mata Atlântica no período 1995 ‑ 2000. Fundação SOS Mata Atlântica/INPE, 2001.
Obs: Os valores absolutos e relativos dos remanescentes florestais nos anos de 1985 e 1990 foram corrigidos a partir da análise das imagens de satélite de 1995, cujo aprimoramento no processamento proporcionou material de maior qualidade para a interpretação visual e assim, apresenta diferenças em relação aos valores divulgados no relatório do Atlas publicado em 1993 (Fundação SOS Mata Atlântica/INPE, 1998).
Neste momento, destaca-se que a região da "Costa Verde" representa um dos maiores núcleos representativos da Mata Atlântica que persistem em todo o estado do Rio de Janeiro (Figura 1). Sendo que nesta região persistem, em diferentes níveis de preservação, além da Floresta propriamente, manguezais e restingas.
Figura 1 - Cobertura florestal remanescente para o estado do Rio de Janeiro durante o período 1990-1995. Em destaque a região da Costa Verde
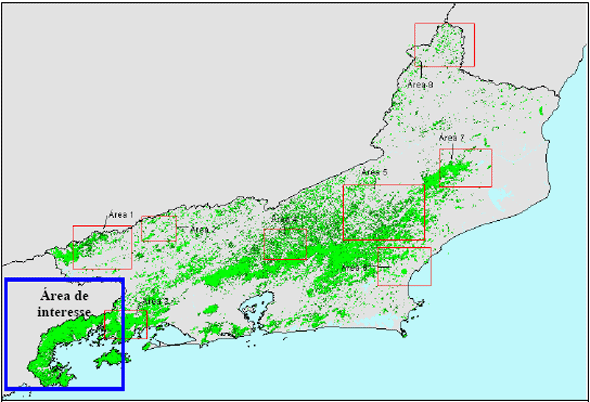
Fonte: Fundação SOS Mata Atlântica/INPE, 1998.
Na Tabela 3 e na Tabela 4, a seguir, apresenta-se a evolução da redução da cobertura vegetal dos remanescentes florestais e dos mais representativos ecossistemas associados (restinga e mangue) pertencentes ao domínio Mata Atlântica no estado do Rio de Janeiro.
Tabela 3 - Evolução da perda de extensão superficial dos remanescentes florestais da Mata Atlântica e seus ecossistemas associados no estado do Rio de Janeiro.
Clique aqui para visualizar a tabela 3
Tabela 4 - Evolução dos Remanescentes Florestais e Ecossistemas Associados da Mata Atlântica no estado do Rio de Janeiro no Período 1995-2000 (SOS Mata Atlântica, 2001).
Classes de mapeamento |
19951 |
20002 |
Desmatamento3 |
| hectares |
%* |
hectares |
%* |
hectares |
%** |
| Remanescentes florestais |
738.402 |
16,82 |
734.629 |
16,73 |
3.773 |
0,51 |
| Restinga |
40.766 |
0,93 |
40.272 |
0,92 |
494 |
1,2 |
| Mangue |
9.865 |
0,23 |
9.610 |
0,22 |
255 |
2,6 |
* em relação à área avaliada do Estado equivalente a 100% / ** em relação aos remanescentes florestais de 1995
Juntamente com outras formações do mesmo gênero, presentes em outros países, a Floresta Atlântica, enquadra-se no que se convencionou chamar de florestas tropicais, bioma terrestre com a maior diversidade biológica do planeta, onde são endêmicas mais de 50% das espécies arbóreas da Terra e diversificada fauna.
É provável que a Floresta Atlântica tenha sido o primeiro bioma florestal a colonizar a Serra do Mar no final do Cretáceo e início do Terciário, o que se deu à cerca de 70 milhões de anos atrás.
A Floresta Atlântica, o ecossistema florestal, já recebeu diversas denominações:
- Floresta Pluvial (Schimper, 1903);
- Floresta Latifoliada Tropical Úmida de Encosta (Azevedo, 1959);
- Formação da Encosta Atlântica (Magnanini, 1965);
- Floresta Perenifólia Latifoliada Higrófila Costeira (Lima, 1966);
- Floresta Ombrófila Densa (Ellenberg & Muller - Dombois, 1965/6);
- Floresta Pluvial Atlântica (Rizzini, 1979).
A designação atual para estas florestas as enquadra na categoria da Floresta Ombrófila Densa, termo proposto por Ellenberg E Mueller -Dombois (1965/6), sendo que o termo Ombrófila (de origem grega), substitui Pluvial (de origem latina) adotado por Rizzini (1979), ambas com o mesmo significado “amigo das chuvas”.
Este tipo de vegetação é caracterizado por fanerófitas (plantas lenhosas com as gemas e brotos de crescimento protegidos, situados acima de 0,25 m do solo, alcançando até 50 m de altura), dominados pelas subformas de vida macro (de 30 a 50 m de altura) e mesofanerófitas (de 20 a 30 m de altura), além de lianas lenhosas e epífitas em abundância que o diferenciam de outras classes de formações. Porém, sua característica ecológica principal reside nos ambientes ombrófilos que marcam muito bem a região florística vegetal.
Este bioma ocupa a área tropical, dominada por temperaturas elevadas (médias de 25oC), valores de precipitação elevados, com período seco anual variando de 0 a 60 dias, e caracteriza-se pela presença de fanerófitas perenifoliadas, com brotos foliares geralmente desprovidos de proteção contra a seca.
Os ambientes ocupados pela Floresta Ombrófila Densa, apresentam chuvas bem distribuídas, com médias anuais em torno e acima de 1500 mm, havendo estações sem seca ou mesmo com grande disponibilidade de umidade.
A subdivisão da Floresta Ombrófila Densa realizada pelo RADAMBRASIL (1983), baseou-se principalmente na distribuição por altitude, sendo reconhecidas para a região da Costa Verde a:
Floresta das Terras Baixas
Abrange os ambientes situados entre cerca de 5 m acima do nível do mar e a altitude de 50 m. A Floresta das Terras Baixas caracteriza-se pela sucessão desde a fase pioneira dos locais salobros com vegetação higrófila de Eleocharis, Typha, Cyperus até a fase fanerófita de Ficus, Tabebuia e Arecastrum, ainda em áreas úmidas, mas com água doce, com uma fase intermediária de Callophillum, Tapirira e Geonoma (RADAMBRASIL, 1983). Abrangeria, portanto nesta fase pioneira o que é denominado, Floresta de pântano ou Mata alagadiça, que apresenta representantes preservados na Reserva Biológica Estadual da Praia do Sul, na Ilha Grande (Maciel et al., 1984).
Na parte bem drenada, ocorrem os grupamentos mais estáveis, da Floresta de Terras Baixas, que além de conservarem alguns componentes das fases anteriores, apresentam um estrato dominante de cerca de 25 m de altura, com tanheiro (Alchornea triplinervia), sangue-de-drago (Croton sp.), figueira do brejo (Ficus organensis), ipê-.do-brejo (Tabebuia sp.); um estrato dominado de cerca de 20 m de altura com bicuiba (Virola sp.), pindaíba (Xilopia sp.), freijó (Cordia sp.), pau jacaré (Piptadenia gonoacantha), angico-branco (Parapiptadenia sp.); um estrato intermediário de aproximadamente 10 m de altura com ingá (Inga sp.), Posoqueria sp., candiúba (Trema micrantha) e um estrato inferior onde predomina a bananeira-do-mato (Heliconia sp.), em meio à ocorrência generalizada de indivíduos das famílias botânicas Palmae, Bromeliaceae, Orquidaceae e Pteridophytae. A estrutura fanerófita da formação apresenta ainda lianas e epífitas em abundância (RADAMBRASIL, 1983).
Floresta Submontana
Ocorre na faixa de altitude entre 50 e 500 m, em áreas dissecadas da Serra do Mar, das serras litorâneas e dos maciços isolados, sobre rochas do embasamento cristalino e rochas alcalinas.
Agrupamentos remanescentes desta formação florestal são encontrados no Estado do Rio nas encostas da Serra da Bocaina, da Serra da Tiririca, da Serra da Tijuca e da Serra da Pedra Branca, maciços isolados como o do Tinguá, Mendanha, Gericinó, Serra do Garcia e nas encostas interiorizadas da Serra dos Órgãos e Serra das Araras, ainda sob influência da massa tropical marítima.
Apresentando sempre algum grau de intervenção antrópica podem ser caracterizadas por possuir estrutura fanerófita, com ocorrência de caméfitas, epífitas (dentre elas as orquídeas e bromélias) e lianas e a presença de um estrato de até 25-30 m de altura com murici ou pau-de-tucano (Vochysia tucanorum); baguaçu (Talauma organensis); faveira (Parkia sp.); jacatirão (Miconia theaezans); Plathymenia foliolosa, Alchornea triplinervia, canelas (Nectandra sp. e Ocotea sp.), sangue-de-drago (Croton sp.); pela ocorrência do cada vez mais escasso palmito (Euterpe edulis);Geonoma sp. e também do ameaçado de extinção xaxim. Existe alta freqüência de embaúbas (Cecropia sp.) e é comum encontrarem-se no estrato inferior plântulas, indicando a regeneração de espécies do estrato dominante, algumas Criptófitas e uma poucas caméfitas das famílias Rubiaceae, Myrtaceaee Melastomataceae(Radambrasil, 1983).
De uma forma geral, a vegetação remanescente da Formação Submontana da Floresta Ombrófila Densa, está hoje abrigada em áreas de preservação permanente, ou fazem parte de parques ou reservas equivalentes, pois se situam em encostas com declive acentuado e estão associadas a nascentes de mananciais.
Floresta Montana
Os ambientes da Formação Montana da Floresta Ombrófila Densa, na latitude da Baía da Ilha Grande, ocupa faixa de altitudes entre 500 a 1500 m, sobre rochas do embasamento cristalino e rochas alcalinas.
A Floresta Montana ocupa locais de relevo fortemente dissecado, íngreme e de acesso difícil, permitindo que seja parcialmente poupada. No estado do Rio de Janeiro, os remanescentes desta formação florestal podem ser encontrados principalmente no Parque Nacional da Serra dos Órgãos e na Serra da Bocaina.
A vegetação da Floresta Ombrófila Densa Montana se caracteriza por apresentar um estrato dominante com altura até cerca de 25 m, com espécies tais como: Vochysia laurifolia, Talauma organensis, Cariniana excelsa, Clethra brasiliensis, Ocotea sp., Nectandra sp., dentre as macrofanerófitas, e um estrato dominado de meso e nanofanerófitas de diversas espécies das famílias Rubiaceae, Myrtaceae e Melastomaceae, e mais a presença generalizada de Palmae(palmito, guaricanga, tucum), Pteridophytae(samambaia e xaxim), Bromeliaceae, além degrande quantidade de outras epífitas e lianas.
No estado do Rio de Janeiro os remanescentes bem preservados da Floresta Ombrófila Densa Montana, ocorrem na Reserva Ecológica de Macaé de Cima, excelentemente descrita fitofisionomicamente por Bruni & Lima (1994): “O denso folhiço recobre o solo da floresta e por entre a umidade constante dos troncos e rochas afloram inúmeras ervas como as aráceas que se destacam no cenário monocromático da floresta, ora por suas inflorescências pálidas ora por suas espatas coloridas, entre elas, Anthurium harrisii, A. theresiopolitanum, Philodendron edimundoi, P. ochrostemon e Xanthosoma sagittifolium”. Para a região da Costa Verde os poucos trabalhos produzidos concentram-se na Ilha Grande.
As samambaias são sempre abundantes quando a umidade é mais elevada, seja em pequenas depressões, falhas de rochas ou margens dos rios e córregos, onde suas folhas testemunham o gotejar contínuo das águas límpidas sobre as lâminas foliares que os margeiam. Ilustrando, cita-se, Aspleniurn rnucronaturn, Blechnum macahense, Lindsaea bifida, Stigmatopteris caudata, Gleichenia squamosa e Huperzia friburgensis, entre outras. Espécies como Peperonia corcovadensis destacam-se pela delicadeza de suas folhagens crassas por sobre rochas que afloram no caminho.
A intensidade de cores é acentuada ainda nas flores das gesneriáceas tão abundantes na Reserva de Macaé de Cima. Igualmente presentes no sub-bosque, o vermelho, o coral, o laranja de suas flores destacam-se revelando inúmeras espécies como, Besleria fasciculata, B. macahensis, B. melancholica, Codonanthe cordifolia e Sinningia cooperi. Conquistando ainda mais o espaço aéreo da mata surgem as epífitas Codonanthe grassilis, Nematanthus crassifolius, N. hirtellus.
No que tange as epífitas a natureza da serra de Macaé, é prodigiosa. Muitos são os representantes das bromeliáceas, cujos tanques vigorosos ocupam os troncos e ramos num gradiente vertical apreciável no interior da mata. A ilustrar tal riqueza encontram-se Quesnelia lateralis, Q. liboniana, Pitcairnia carinata, Tillandsia aeris-incola, Vriesia haematina, V. hydrophora e Wittrockia bragarurn.
Tão diversas surgem também as orquidáceas com nuances do violáceo, vinoso e amarelo, como Beadlea warmingii, Chytroglossa marileoniae, Dichaea pendula, Epidendrum addae, E. paranaense, E. xanthinum, Gomesa recurva, Maxillaria cerifera, M. ubatubana, Miltonia cuneata, Oncidium uniflorum, Pabstia jugosa, P. triptera, Phymatidium delicatum, P. tillandsoides, Pleurothalli strifïda, Prescottia epiphyta, Rodrigueziopsis microphyta, Scaphyglottis modesta, Sophronitis aff. mantiquerae e Zygopetalum crinitum, entre outras.
Cabe ressaltar ainda, entre as epífitas, algumas espécies de samambaias como Elaphoglossum alpestre, E. burchellii, E. ornatum, Hymenophyllum rufum e Huperzia heterocarpa e representantes das cactáceas como Hatiora salicornioides, Rhipsalis capilliformes, R. clauata, R. houlletiana, R. itrigona e Schlumbergera truncata esta última conhecida como flor-de-maio.
O sub-bosque é denso e diverso, seus componentes apresentam caules de pequeno calibre, muitos deles ramificando a menos de 1 m do solo, quase sempre recobertos pelas inúmeras briófitas que ai ocorrem. Nos trechos em que a luminosidade passa com maior facilidade, são comuns as presenças de piperáceas como Piper jaequilaterum, P. caldense, P. gaudichaudianum, P. lhotzkianum, P. malacophytlum, P. richardiifolium, quase sempre com suas folhas danificadas pela predação de herbívoros.
As taquaras como Chusquea capitata, C. capituliflora, Guadua tagoara e Merostachys lischeriana, cujos ramos vigorosos abraçam as copas dos arbustos, dificultam a disputa pela luz por parte das outras espécies típicas deste estrato assim como também a dos exemplares jovens de elementos do dossel que começam a chegar na formação da floresta.
Neste sentido, são comuns os jovens de Attalea dubia e Lytocarium insignis nestas áreas mais alteradas. É neste estrato que as mirtáceas, rubiáceas e melastomatáceas são mais freqüentes, encontrando-se seus representantes nas mais diversas alturas e formas.
Floresta Alto Montana
Os ambientes da Formação Florestal Alto Montana, na latitude da baía da Ilha Grande, ocupam faixa de altitude acima do nível de 1500 m, sobre rochas do embasamento cristalino. Por estarem localizadas nas partes mais elevadas são freqüentemente envoltas por densas nuvens durante dias seguidos. É uma típica mata úmida de neblina.
De um modo geral, seus solos são rasos, havendo o desenvolvimento de Solos Litólicos e Cambissolos. O porte da vegetação em função do solo raso, elevada altitude e temperaturas baixas, apresenta quase sempre uma estrutura de nano e microfanerófitas, cuja altura pode variar em torno de 5 a 10 m.
Embora composta por espécies endêmicas, revelando um antigo isolamento, a composição florística desta formação é representada por famílias de dispersão universal, cujos gêneros mais comuns são: Drymis, Clethra, Meimmannia, Rapanea, Hexachlamys, Marliera, Roupala e Miconia. Ocorrem também Bromeliaceaedos gêneros Vriesia (inclusive a magnífica Vriesia imperialis), Aechmea e Nidularium; Cyperaceaerepresentadas pelo gênero Cyperus; a Gramineae Chusquea mimosa e muitas Pteridófitas que recobrem o terreno (RADAMBRASIL 1983). Como vegetação endêmica mais expressiva, ocorrem no Parque Nacional da Serra dos Órgãos; Mimosa glaziovii, Drosena villosa, Begonia edrnundoi e Hidrocatyle quinquiloba (Ibama, 1994).
Em função das baixas temperaturas, freqüentemente com médias abaixo de 15oC, é comum a vegetação desta formação florestal se apresentar com formas xerofíticas, caracterizadas pelos troncos e galhos finos, casca rugosa, folhas ericóides, pequenas, coriáceas ou carnosas, e brotos terminais protegidos. Normalmente há grande incidência de epífitas e líquens que por sua vez indicam a existência de altos teores de umidade relativa do ar no ambiente local (Radambrasil, 1983).
Por se situar em área de relevo acidentado e de difícil acesso, este tipo florestal foi bastante poupado, não estando, no entanto, imune à ação antrópica.
Apesar de sua importância histórica, pois é das mais antigas áreas de ocupação no litoral, não encontramos uma quantidade muito extensa de trabalhos sobre as formações vegetacionais em Angra dos Reis, como seria de se esperar. No entanto, podemos adicionar informações através da consulta a trabalhos feitos em localidades próximas, como Parati e principalmente no litoral Norte do estado de São Paulo, para ter uma idéia da vegetação da região do entorno do município de Angra dos Reis.
A região de Angra dos Reis apresenta pluviosidade descontínua espacial e temporalmente, não ocorrendo, no entanto, real escassez de água nem durante o inverno. Destaca-se que as intensas chuvas durante o verão geram problemas como enchentes e assoreamentos, que estão associados ao desmatamento no platô da Serra da Bocaina (RJ/SP) e na planície fluvio-marinha.
Os ambientes da orla litorânea da região sofreram grande alteração antrópica (e.g. desmatamento, erosão e impermeabilização dos solos). Existem dados, em vários relatórios técnicos que evidenciam que as formações vegetais entre 1966 e 1987, um aumento da área relativa de gramíneas, associado muitas vezes, ao desmatamento no platô da Bocaina, sobre as planícies e porções inferiores das encostas.
A área das formações florestadas, no entanto, não sofreu alteração significativa no período estudado, possivelmente em razão da regeneração natural de outras áreas, que pode estar refletida na redução na área relativa de formações iniciais, ou de “capoeiras”.
Registram também um aumento expressivo das áreas urbanas, sobretudo pela expansão da cidade de Angra ao longo da BR-101 (Rio-Santos). Os autores citam o surgimento de concentrações na planície no baixo Rio Bracuí, baixo Rio Mambucaba (Perequê) e enseada do Frade. A rodovia Rio-Santos criou a especulação imobiliária ao longo da estrada, com a construção de complexos hoteleiros, condomínios e marinas.
Associada à construção da estrada, houve uma expansão do turismo, instalação de loteamentos para casas de veraneio, assim como para as residências de funcionários de Furnas (Mambucaba, Itaorna e Praia Brava) e pequena rede de serviços. Conforme os autores, a expansão das áreas urbanas se deveu em detrimento de áreas de restingas, manguezais e brejos, que sofreram uma redução de área total de 46,0 km2 em 1966 para 21,9 km2 em 1987.
Conforme relatório técnico sobre a vegetação da Bacia do Rio Paca-Grande (Farag et al., 1997), no platô da Serra da Bocaina, é mais florestada que a bacia do Rio Bananal, com 83,3% de cobertura de formações florestais de pioneiras até tardias (clímax). Segundo os autores, as formações pioneiras e secundárias são resultado do abandono de áreas de pastagem e de exploração de carvão. As formações tardias ocupam áreas de difícil acesso ou impróprias para o uso, ou ainda em áreas para a conservação de mananciais e ao longo das drenagens. No levantamento florístico realizado, os autores encontraram 140 espécies pertencentes a 44 famílias.
Marques et al. (1997), em estudo na APA do Cairuçu, próximo à cidade de Parati, com uma área de 33.000 ha e 63 ilhas e relevo acentuado (até 1320 m.s.m.), citam que esta é ocupada por Floresta Ombrófila Densa, apresentando áreas que sofreram corte seletivo e áreas de culturas e pastagens, onde o desmatamento foi intenso.
Os autores encontraram um total de 972 espécies de plantas com flores, além de pteridófitas (samambaias) (115 spp.), musgos (124 spp.) e algas (117 spp.). A floresta possui altura de cerca de 20 a 25 m com emergentes de 30 a 35 m. Os autores assinalam a grande quantidade de plântulas de palmito (Euterpe edulis).
Eles mencionam também que a vegetação mais preservada foi observada à distância nos vales profundos e encostas de difícil acesso e alguns morros. Citam que até 300-350 m.s.m. a população local utiliza a mata como fonte de madeira, e que de um modo geral são pequenas as áreas de mata preservada.
Em dissertação de mestrado sobre um trecho de Mata Atlântica de encosta em Ubatuba - SP, Silva (1980) amostrou indivíduos maiores de 10 cm de DAP e encontrou índices de diversidade comparáveis a valores de matas de terra-firme na Amazônia. As famílias botânicas com maior riqueza de espécies foram Myrtaceae, Lauraceae, Leguminosae, Rubiaceae, Euphorbiaceae e Sapotaceae.
Este autor menciona que a mata atlântica após a subida da Serra do Mar cedeu lugar a uma vegetação campestre, enquanto aquela ficou restrita a algumas manchas isoladas, particularmente nas encostas.
Tabarelli & Mantovani (1999) em trabalho sobre a diversidade de florestas de encosta (i.e. Floresta Ombrófila Densa) no estado de São Paulo (e.g. Juréia, Ilha do Cardoso, Cubatão, Picinguaba, Ubatuba e Salesópolis) mostram que as famílias com maior riqueza de espécies são Myrtaceae, Leguminosae, Rubiaceae, Lauraceae, Melastomataceae e Euphorbiaceae, todas com mais de 15 espécies.
Segundo estes autores, as referidas formações têm diversidade menor de espécies arbóreas que outras áreas de florestas neotropicais, sobretudo em floretas amazônicas de baixada, destacando, no entanto, os altos níveis de endemismo das florestas atlânticas de encosta. Grande parte da riqueza de espécies nestas formações se deve às famílias Melastomataceae, Rubiaceae, Myrsinaceae, Myrtaceae, Monimiaceae, Arecaceae e Flacourtiaceae.
Sanchez et al. (1999) estudando a vegetação do Rio da Fazenda em Picinguaba, município de Ubatuba (SP), medindo indivíduos maiores que 6,36 cm de DAP em parcelas com 0,4 ha, encontraram maior riqueza para Myrtaceae, seguida de Rubiaceae, Arecaceae, Sapotaceae e Cyatheaceae. Euterpe edulis, o palmito, foi a espécie com maior número de indivíduos (11,7% do total). Os autores sugerem que a alta diversidade encontrada se deve à heterogeneidade do ambiente em relação à luminosidade e dos solos.
Tabarelli & Mantovani (1997; 1998) amostraram a vegetação colonizadora de clareiras pequenas, no Parque Estadual da Serra da Cantareira (SP) e encontraram que apenas 37,5% são espécies intolerantes à sombra, sobretudo grandes pioneiras.
Os autores mediram indivíduos maiores que 10 cm de CAP (circunferência à altura do peito) em 0,2 ha de mata adjacente às clareiras que teve 56,43% (N=13) das espécies do subosque da mata típicas desse ambiente, enquanto que o restante é constituído de espécies típicas de dossel.
7.1.2.1. Composição Florística e Estrutura Fitossociológica- topo
O conhecimento da composição florística e da estrutura fitossociológica das espécies têm muito a contribuir para a conservação, a recuperação e o manejo deste ecossistema.
Estudos fitossociológicos mostram o papel exercido por cada espécie dentro da comunidade (Rodrigues & Gandolfi, 1998) e também contribuem de forma decisiva na indicação dos estágios sucessionais e para uma melhor avaliação da influência de fatores de clima, solo e ação antrópica nas comunidades vegetais.
Para a realização destes estudos são, basicamente, utilizadas duas metodologias distintas: o método de ponto quadrante Martins, 1979 e o método de parcelas proposto por Mueller - Dombois & Ellemberg, 1974 e utilizado por Fournier & Fournier, 1985; Baitello et al., 1988; Silva et al., 1993; Rizzini et al., 1997 e Borém & Ramos, 2001.
O método de ponto quadrante tem a vantagem de cobrir uma maior área, por isso é considerado um método exploratório e recomendado para o estudo em áreas extensas.
Já no método de parcelas a localização das unidades amostrais é mais fácil, fazendo com que a continuidade dos estudos seja mais simplificada em tempos futuros. Além disso, este método possibilita estudos da regeneração natural e da dinâmica da população através do acompanhamento da natalidade, do crescimento e da mortalidade dos indivíduos presentes nas unidades amostrais. O entendimento destes diferentes processos e estágios é imprescindível para qualquer plano de estudo, manejo e conservação de formações naturais dos ecossistemas.
O presente estudo baseia-se nos dados coletados em amostragens realizadas em áreas selecionadas previamente através de excursão de reconhecimento.
Para o levantamento fitossociológico foi adotado o método de parcelas (Mueller-Dombois & Ellemberg, 1974). A amostragem foi realizada através da instalação de 20 parcelas de 20 x 25 m, totalizando uma área de 10.000 m2 (1,0 ha), para cada ambiente amostrado. Cada parcela foi delimitada por estacas de madeira de 1 m e cercada com cordão de poliéster.
A escolha pelo método de parcelas justifica-se em função do mesmo apresentar melhores vantagens em relação a outros métodos fitossociológicos, e, em permitir correlações mais estreitas com os fatores atuantes na área, além de fornecer melhores subsídios para o entendimento da distribuição espacial das espécies e dos diferentes estágios de desenvolvimento presentes no ambiente.
Como unidade amostral considerou-se todos os indivíduos vivos, lenhosos, com diâmetro do caule ao nível do peito (DAP) ³ 5 cm e altura total (AT) ³ 1 m, que ao nível do solo estivesse dentro da parcela. Indivíduos que apresentaram parte do caule dentro da parcela ou que tocaram por fora as linhas de limites das parcelas foram amostrados. Não foram considerados no tratamento dos dados rebrotos provocados pela ação do fogo ou reprodução vegetativa, subarbustos, palmeiras acaule e bromeliáceas, orquidáceas e cipós/trepadeiras.
Todos os indivíduos selecionados no critério de inclusão foram etiquetados com plaquetas de PVC e numerados em ordem crescente independente da numeração das parcelas. Nas fichas de campo foram anotados os seguintes dados, quando possível:
As medidas de circunferência ao nível do peito foram efetuadas com fita métrica de 1,5 m e as alturas estimadas com o uso de uma vara de alumínio regulável, intercambiável, graduada a cada 0,5 m e perfazendo um total de 9m com a vara toda distendida. Indivíduos maiores que 9 m tiveram as suas alturas estimadas visualmente a partir do ápice da vara.
Foram anotadas também informações complementares das parcelas e de suas imediações como tipo de solo, aspectos de relevo e drenagem, presença de serrapilheira, tipo de dossel, espécies indicadoras que não entraram no tratamento dos dados, vestígios de fogo, dentre outros.
Quando foi possível tomar coordenadas espaciais, estas foram determinadas com o uso de GPS Garmin modelo 45XL.
Em cada área amostrada foram coletadas amostras de todos os exemplares que não eram conhecidos pelos coletores. Para o levantamento florístico, foram coletadas também as espécies que apresentavam material reprodutivo, localizadas nas proximidades das parcelas, desde que atendessem ao critério de inclusão.
As coletas foram feitas com tesoura de poda manual e de poda alta, o material reprodutivo (flores, botões e frutos) e o material vegetativo foram coletados, acondicionados em sacos de coletas e prensados ao final do dia com a elaboração de uma ficha de identificação para cada espécie. O número de exemplares, o acondicionamento, a prensagem e a herborização foram processados conforme Castro (1987), sendo observados todos os cuidados e detalhes indispensáveis à coleta, preservação e herborização.
A identificação do material botânico deu-se previamente no campo através da análise dos indivíduos coletados, sendo posteriormente conferida, complementada e/ou corrigida por morfologia comparada, com outros materiais já identificados. O material coletado em campo foi entregue ao Herbarium Bradeanum (sigla HB), conveniado com a Universidade do Estado do Rio de Janeiro - Uerj, que providenciou sua secagem e preparação para identificação botânica. Todo o material coletado e preparado foi identificado através da metodologia clássica de morfologia comparada, foi feito o uso de chaves de identificação, literatura especializada e comparação com exsicatas, bem como o posterior auxílio de especialistas. Quando não foi possível a identificação do material coletado até o nível de espécie, esta se deu até a categoria taxonômica mais abrangente mais próxima, no caso Gênero ou Família, sendo considerada então como morfoespécie (exemplo: Cordia sp1., Myrtaceae sp1.). Quando não foi possível a identificação sequer ao nível de Família Botânica (14 casos), esta recebeu o código para indeterminada “Indet.”, sendo separadas de acordo com suas características anatômicas em grupos (Indet. sp. 2, Indet. sp.3).
Os dados foram tabulados e elaborados gráficos para facilitar análise em função dos diversos objetivos.

A suficiência da amostragem foi avaliada através da curva de incremento de espécies por área (curva espécie - área), proposta por Mueller-Dombois & Ellemberg, 1974.
De um modo geral, a maior parte da cobertura florestal ocorrente nas adjacências do empreendimento recobre as vertentes atlânticas da Serra do Mar, estando concentrada principalmente em áreas declivosas pouco propícias à ocupação humana e à implantação de culturas.
Originalmente, em consonância ao sistema de classificação proposto por Veloso et al. (1991) e adotado pelo IBGE (1992), a cobertura florestal na região integra parte da região fitoecológica da Floresta Ombrófila Densa. A ocorrência deste tipo de vegetação relaciona-se a fatores climáticos com elevadas temperaturas (médias de 25°C) e uma pluviosidade bem distribuída durante o ano, sem um período biologicamente seco. De um modo geral, caracteriza-se pela cobertura arbórea densa com fanerófitos perenifólios, estruturados em vários estratos e associados a trepadeiras lenhosas e epífitas.
O estado de conservação das florestas locais varia de acordo com o grau de interferência antrópica em cada trecho. Assim sendo, parte da cobertura é atualmente constituída por uma vegetação secundária em estágios diversos de regeneração, ao passo que outras áreas são recobertas por florestas alteradas em sua composição e estrutura originais, tendo em vista a extração seletiva de madeiras a qual foram submetidas ao longo dos anos. Por outro lado, remanescentes florestais em melhor estado de conservação podem ser observados em certos trechos, em geral recobrindo encostas íngremes de difícil acesso, ou sobre terrenos com substrato litólico, desfavoráveis a culturas.
Algumas áreas de encosta são atualmente aproveitadas para atividades agrícolas, onde se destaca o cultivo de banana. Nas matas atingidas pela extração madeireira, a intensidade da alteração varia com a declividade e a dificuldade de acesso. De um modo geral, boa parte dos fragmentos florestais locais encontra-se desprovido de elementos arbóreos de maior calibre e valor comercial, os quais originalmente compunham os estratos superiores da mata.
Outrossim, diversos trechos são revestidos por uma vegetação secundária, que se desenvolveu após a remoção da vegetação original. O processo de sucessão natural envolve a substituição gradativa de espécies adaptadas a cada uma das comunidades sucessionais, e são o reflexo de diversos fatores atuantes, podendo-se citar, entre eles, o tempo de uso e de abandono do solo, e também a forma de manejo aplicada em cada área (desmatamentos, queimadas, etc.).
Nas áreas recentemente abandonadas após o uso da terra, ocorre um processo pioneiro de colonização do solo através do desenvolvimento de gramíneas e plantas invasoras características das fases iniciais de sucessão secundária. O desenvolvimento desta vegetação irá preparar o solo e as condições para o surgimento de espécies arbustivas, formando as popularmente conhecidas “capoeirinhas”, onde predominam espécies de compostas, verbenáceas, malváceas, tiliáceas, labiatas e melastomatáceas, entre outras.
Em fase posterior a esta, havendo continuidade no processo sucessional, surge uma vegetação mais desenvolvida, conhecida popularmente por “capoeira”, caracterizada pelo predomínio de espécies arbustivas e arbóreas em sua maioria pioneiras e secundárias. Em trechos secundários locais é comum a ocorrência de espécies como Tibouchina sp. (quaresmeira), Cecropia glazioui (embaúba-vermelha), Croton sp., Sparattosperma leucanthum (ipê-branco), Cupania oblongifolia (camboatá), Piptadenia gonoacantha (jacaré), Schinus terebinthifolius (aroeira) e Zanthoxylum rhoifolium (maminha-de-porca) entre outras.
Neste levantamento foram inventariadas duas fisionomias da Floresta Ombrófila Densa, que diferem principalmente com relação aos seus estados de conservação. Uma delas está situada na cadeia montanhosa adjacente à CNAAA, sendo dividida em dois trechos, um na vertente onde se situam as torres de comunicação da empresa e a outra na vertente de acesso à chamada praia do Mamede (Figura 2). Esta fisionomia também apresenta indícios da atividade humana, seja pela ausência de exemplares de porte elevado como pela presença de restos de troncos em clareiras em sucessão. Esta fisionomia será chamada de “Floresta 1”.
A outra região amostrada está situada na bacia do córrego praia Brava, área da antiga captação de águas para o condomínio praia Brava, apresenta indícios de extração seletiva de espécies de maior interesse, persistindo, porém exemplares com porte elevado (Figura 2). Foram encontrados pelo menos dois exemplares de limoeiros em meio à mata, indicando pretérita presença humana nesta bacia. Além disso, foram obtidos relatos da presença de habitações nesta região, porém sem definir quantas e onde. Esta fisionomia passa a ser chamada de ”Floresta 2”.
Figura 2 - Vista geral da área onde foram realizados os levantamentos fitossociológicos que fazem parte deste relatório. A: Floresta 1; B: Floresta 2.

- Floresta 1 (Área das torres de comunicação e encosta da praia do Mamede)
Localiza-se em trechos de matas, de encosta, alteradas, cuja vegetação foi afetada pela extração de madeira, cultivo de banana e também por obras de construção de estrada no caso da área das torres de comunicação. Apresentam um dossel mais ou menos contínuo, onde sobressaem espécies arbóreas como Ficus (e.g. Ficus insipida, F. arpazuza), Virola cf. gardneri, Alchornea triplinervia, Hyeronima alchorneoides, Quararibea turbinata, Cordia sp., Sloanea sp. e Cedrela cf. odorata, entre outras.
O interior destas matas mostra-se em geral denso e marcado pela intensa regeneração de indivíduos arbóreos dos estratos superiores. Arbustos e árvores comuns na submata são ilustrados por gêneros como Urera, Guarea, Siparuna, Guapira (G. opposita), Miconia, Trichilia, Psychotria e Piper. Entre as palmeiras aí encontradas destaca-se o iri (Astrocaryum aculeatissimum). É comum a ocorrência de trepadeiras lenhosas (lianas) representadas por gêneros como Smilax, Serjania, Mikania, Adenocalymma, Arrabidaea, Acacia, Bauhinia e Peltastes (P. peltatus). Ervas são por sua vez exemplificadas por Heliconia cf. angusta, Anemia phyllitidis, Costus spiralis e Pothomorphe umbellata.
Entre as epífitas e hemiepífitas mais comuns sobressaem espécies da família Bromeliaceae (e.g. Vriesea spp., Nidularium spp.), Cactaceae (Rhipsalis spp.) e Araceae (Anthurium spp., Philodendron spp. e Monstera adansonii).
Em particular na área da praia do Mamede foram observados indícios da extração de palmito e também armadilhas de espera (trepeiro).
- Floresta 2 (Área da antiga captação de águas da praia Brava)
As parcelas foram implantadas na bacia do córrego praia Brava e corresponde a um trecho de floresta em bom estado de conservação. Neste ambiente, árvores desenvolvidas compõem um dossel em torno de 15 a 20m de altura. O estrato superior é diversificado, exibindo espécies das famílias Myrtaceae, Lauraceae, Annonaceae, Leguminosae, Cecropiaceae (Pourouma guianensis) e Araliaceae (Didymopanax cf. anomalus), entre diversas outras.
A densa cobertura arbórea confere ao interior desta mata um ambiente sombrio e úmido, e o estrato inferior aberto facilita a passagem. São comuns as espécies arbustivas e arbóreas de famílias como Monimiaceae (Siparuna sp., Mollinedia sp.),Melastomataceae e Rubiaceae (Psychotria spp.), destacando-se ainda espécies de palmeiras (Geonoma sp., Astrocaryum aculeatissimum ) e pteridófitas arborescentes (Cyathea spp.).
Entre as sinúsias dependentes de suporte, destacam-se diversas trepadeiras lenhosas ou lianas, várias das quais bem desenvolvidas, que se desenvolvem apoiando-se em árvores hospedeiras de forma a atingir o dossel da mata. São ilustradas por espécies das famílias Bignoniaceae, Leguminosae e Menisperpaceae de bignoniáceas, leguminosas, menispermáceas (Chondodendron platyphyllum) e Sapidaceae (Paullinia spp., Serjania spp.), entre outras.
Este trecho é ainda marcado pela abundância de epífitas, representadas por espécies de bromélias (Nidularium spp., Aechmea sp., Vriesea spp., Quesnelia spp.), Araceae (Anthurium spp.), Cactaceae (Rhipsalis spp.), Piperaceae (Peperomia spp.) e Orquydaceae, entre outras. Hemiepífitas são ilustradas por Moraceae (Ficus spp.) e Araceae (Philodendron spp., Monstera adansonii) e Solanaceae (Dyssochroma viridiflora).
Os estratos inferiores neste trecho são formados por diversos elementos arbustivos e herbáceos, destacando-se, entre os primeiros, diversas espécies de Monimiaceae, Myrtaceae e Rubiaceae. Entre as ervas, são freqüentes espécies de Moraceae (Dorstenia spp.), begoniáceas (Begonia spp.), Marantaceae, Heliconiaceae (Heliconia spp.) e Zingiberaceae(Renealmia petasites), entre outras.
Em trechos próximos às margens do córrego, o substrato pedregoso favorece a ocorrência de espécies rupícolas que sobrevivem no interior da mata, onde sobressaem diversas espécies de bromélias (Vriesea spp., Nidularium spp., Neoregelia spp.) e Araceae.
Para esta região o levantamento executado pela CNAAA inventariou diversas espécies entre arbóreas, epífitas, etc, várias destas não observadas nas áreas de estudo.
Foram amostrados no total 2.328 indivíduos, distribuídos em 51 Famílias, 116 gêneros e 236 espécies/morfoespécies para uma área total de 2,0 ha de Floresta Ombrófila Densa.
A listagem com as famílias ocorrentes nas duas áreas amostradas e o número de espécies por família se encontram na Tabela 5.
Tabela 5 - Listagem de famílias botânicas encontradas e número de espécies, total e por formação inventariada.
Clique aqui para visualizar a tabela 5
Nos estudos realizados por NATRONTEC (1998), na floresta em frente a CNAAA, foram obtidos 313 indivíduos arbóreos, distribuídos dentre 114 espécies, 64 gêneros e 38 morfoespécies para uma área amostral de 0,2ha. À partir dos dados totais levantados foi estimada uma densidade média de 1.565 ind.ha-1 de mata para um CAP ³ 10 cm (DAP ³ 3,18 cm).
A listagem das famílias e espécies encontradas de acordo com a formação (Floresta 1 ou Floresta 2) onde foram encontradas é apresentada no Anexo 1. Os parâmetros fitossociológicos, por sua vez, são apresentados separadamente para a Floresta 1 (Anexo 2) e para a Floresta 2 (Anexo 3).
Considerando as duas formações abrangidas neste estudo, foram amostrados, para 1,0 ha da Floresta 1, 840 indivíduos, distribuídos em 39 famílias, 70 gêneros e 104 espécies / morfoespécies. Para 1,0 ha de Floresta 2 foram obtidos 1.488 indivíduos, distribuídos em 47 famílias, 101 gêneros e 205 espécies / morfoespécies. Com relação à similaridade entre as duas áreas florestais amostradas, foram obtidas 35 Famílias comuns, concentrando 189 e 100 espécies / morfoespécies respectivamente.
Com relação à suficiência amostral, foram traçadas as curvas do coletor para as duas formações, sendo exibida na Figura 3. Ambas as curvas indicam não ter sido alcançada a estabilidade na curva, o que indica a necessidade de aumentar mais as áreas amostradas. Neste momento vale ressaltar que as duas áreas onde foram realizados os inventários não constituem florestas primárias, apresentando indícios de interferência antrópica, o que pode levar a este tipo de curva já que a floresta está alterada e existem grandes mudanças de local para local. Ao mesmo tempo, deve-se salientar também que além desta influência, a declividade do terreno também contribui para a queda de árvores, implicando na formação de mosaicos onde existem manchas em diferentes estágios sucessionais lado a lado (Guedes-Bruni et al., 1997; Mantovani et al., 1990).
Figura 3 - Curva do coletor traçada para a área amostrada em Floresta 1 e Floresta 2.
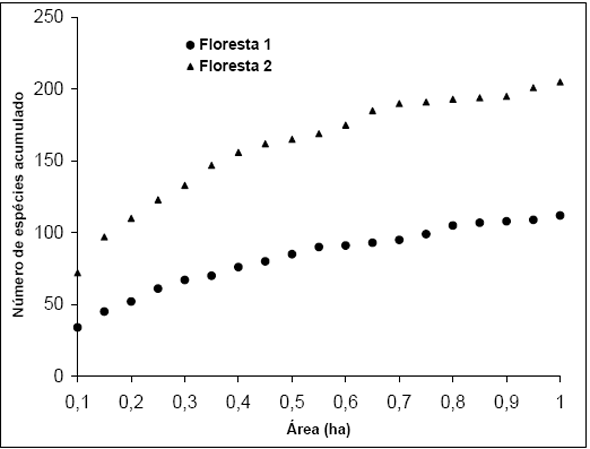
Com relação às Famílias exclusivas para estas áreas, para a Floresta 1 foram obtidas 12 famílias com um total de 16 espécies/morfoespécies, enquanto para a Floresta 2 foram obtidas 4 famílias exclusivas, com 4 espécies/morfoespécies.
O material não identificado correspondeu a 16 exemplares, sendo que oito destes, ou 1%, em Floresta 2 e sete deste, ou 2%, em Floresta 1. Dentre estes existem dois indivíduos, um em cada formação amostrada totalmente desprovidos de folhas na ocasião do inventário.
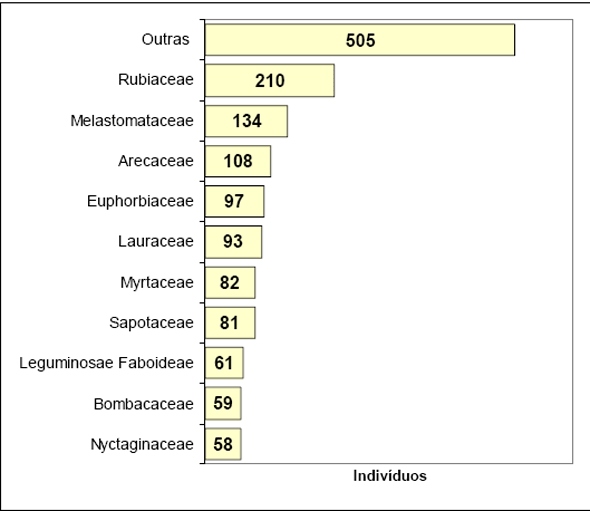
As famílias com maior número de espécies em Floresta 2, foram Myrtaceae (25), Rubiaceae (21), Lauraceae (18), Sapotaceae (13), Leguminosae Faboideae (11), Melastomataceae (9), Euphorbiaceae (9), Moraceae (8), Monimiaceae (7) e Meliaceae (7).
Para a Floresta 1, as famílias com maior número de espécies foram Euphorbiaceae (10), Rubiaceae (9), Myrtaceae (7), Leguminosae Faboideae (7), Moraceae (5), Lauraceae (5), e Piperaceae, Monimiaceae, Melastomataceae, Leguminosae Mimosoideae com 4 espécies cada.
As 10 espécies com maior número de indivíduos para Floresta 2 totalizaram 983 ind. (66%), enquanto que para Floresta 1 obteve-se 556 ind (66%) (Figura 4 e Figura 5). No caso da primeira floresta 44% dos indivíduos são distribuídos entre 37 famílias, enquanto que para a outra temos 29 famílias. Considerando a família Leguminosae sem suas subdivisões, estes valores mudam para 1036 ind. (70%) e 592 ind. (70%) para Floresta 2 e Submontana, respectivamente.
Segundo o relatório elaborado pela NATRONTEC para Angra 2 (1998) as Famílias Botânicas que mais se destacaram quanto ao número de indivíduos foram: Melastomataceae (57); Rubiaceae (25); Euphorbiaceae (20); Leguminosae (18); Arecaceae (=Palmae) (16); Myrtaceae (14) e Monimiaceae (11), que responderam por mais de 50%do total de indivíduos. Sendo que várias destas mesmas espécies também apresentaram destaque quanto ao número de espécies. Assim, temos: Rubiaceae (10); Myrtaceae (8); Melastomataceae (7); seguidas por Euphorbiaceae e Lauraceae (6), Leguminosae (5); o que equivale a 36,5% do total de espécies.
Para a Floresta 2 o maior número de indivíduos foi obtido para a família Rubiaceae (210 ind.) distribuídos em 21 espécies. Já para a Floresta 1 o maior número de indivíduos ficou com a família Lauraceae (97 ind.) que apresentou somente 5 espécies. Nesta última área o maior número de espécies ficou com a família Euphorbiaceae, que apresentou o segundo maior número de indivíduos.
Figura 4 - Listagem das 10 famílias com maior número de indivíduos para Floresta 1.
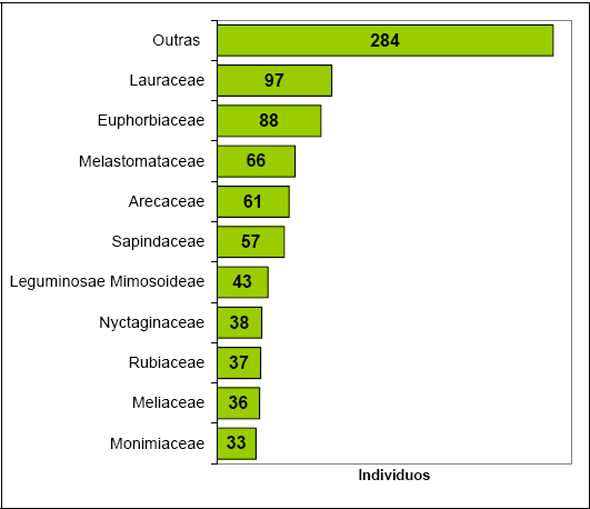
Figura 5 - Listagem das 10 famílias com maior número de indivíduos para Floresta 2.
Com relação à composição específica, para Floresta 1 as espécies que mais se destacaram quanto ao Valor de Importância foram Hyeronima alchorneoides (Euphorbiaceae), Cupania oblongifolia (Sapindaceae), Guapira opposita (Nyctaginaceae), Miconia cinnamomifolia (Melastomataceae), Piptadenia gonoacantha (Leguminosae Mimosoideae), Ficus insipida (Moraceae), Euterpe edulis (Arecaceae), Ocotea puberula (Lauraceae), Cabralea canjerana ssp. canjerana (Meliaceae) e Nectandra leucantha(Lauraceae).
O maior destaque obtido por H. alchorneoides foi devido ao seu elevado valor de área basal, o que a coloca bem à frente da segunda espécie (C. oblongifolia). Além da área basal outra variável que contribuiu foi a maior freqüência absoluta. H. alchorneoides é uma espécie de ampla distribuição e que pode ser encontrada tanto em florestas primárias quanto secundárias. C. oblongifolia também é uma espécie que pode ser encontrada tanto em florestas primárias quanto secundárias, porém em área declivosas e solos férteis. Tanto G. opposita, M. cinnamomifolia quanto P. gonoacantha são espécies heliófitas, comumente encontradas em formações secundárias. A espécie O. puberula também é típica de ambientes secundários, porém sendo rara em ambientes primários. As espécies mais comumente encontradas em florestas mais preservadas, porém não sendo exclusivas destes, são C. canjerana e N. leucantha. De um modo geral, as 10 espécies que apresentaram maiores valores de importância, apresentaram pequenas freqüências absolutas, porém seus valores de área basal foram superiores aos das espécies da Floresta 2. As espécies C. oblongifolia, C. canjerana e E. edulis aparecem entre as mais citadas entre os levantamento em área de Floresta Atlântica.
Com relação à composição específica, para Floresta 2, nota-se que as espécies que mais se destacaram com relação ao Valor de Importância foram Bathysa australis (Rubiaceae), Guapira opposita (Nyctaginaceae), Eriotheca pentaphylla (Bombacaceae), Hyeronima alchorneoides (Euphorbiaceae), Miconia prasina (Melastomataceae), Astrocaryum aculeatissimum (Arecaceae), Vernonia discolor (Asteraceae), Chrysophyllum flexuosum (Sapotaceae), Euterpe edulis (Arecaceae) e Coussarea meridionalis var. porophylla (Rubiaceae).
A espécie B. australis obteve sua maior colocação em função de, principalmente, seu elevado número de indivíduos e sua ampla distribuição nas parcelas amostradas. Esta espécie é heliófita, sendo comumente encontrada em ambientes secundários. Das dez espécies de maior Valor de Importância, seis apresentaram valores de freqüência absoluta superiores a 0,8, sendo o máximo de 1,0. Com relação às espécies que se destacaram, temos que são todas de ambientes secundários.
Em levantamento anteriormente realizado na região de floresta em frente à CNAAA (NATRONTEC, 1998) os maiores Valores de Importância foram obtidos para as Famílias botânicas Euphorbiaceae, Melastomataceae, Rubiaceae, Myristicaceae, Leg - Mimosoideae, Myrcinaceae, Arecaceae, Lauraceae. Com relação à composição específica, mereceram destaque as espécies H. alchorneoides (Euphorbiaceae); B. meridionalis (Rubiaceae); V. oleifera (Meristiceae); I. edulis (Leg - Mimosoideae); M. rimalis, T. canescens e M. aureoides (Melastomataceae); R. ferruginea (Myrcinaceae); E. edulis (Arecaceae); C. moschata (Lauraceae).
7.1.2.2. Diversidade de Espécies- topo
Para a área total amostrada (1,0 ha) em Floresta 2 a densidade foi de 1.488 ind.ha-1, com índice de diversidade de Shannon-Weaver de 2,03 e equabilidade de Pielou de 0,87. Para a Floresta 1 foi obtida uma densidade de 840 ind.ha-1, com índice de diversidade de Shannon-Weaver de 1,77 e equabilidade de Pielou de 0,86. Para fins de comparação pode-se citar Silva (1980) que realizou inventário na região de Ubatuba (SP) e, utilizando o método dos Quadrantes, obteve 123 espécies (DAP ³ 10cm) e para o índice de diversidade de Shannon-Weaver o valor de 3,49. Outro trabalho a ser mencionado foi o de Guedes-Bruni et al. (1996; In: Guedes - Bruni et al. 1997) onde foram obtidos 189 espécies (método das parcelas; DAP ³ 5cm) e para o índice de diversidade de Shannon-Weaver o valor de 4,05. Certamente a adoção de metodologias e limites de DAP distintos dificultam as comparações, mas apesar disto os valores obtidos ficam próximos aos citados na bibliografia especializada.
Estudos realizados em Floresta Ombrófila Densa, tais como em Macaé de Cima, RJ, apontam 2.091 ind.ha-1 para uma área em bom estado de conservação (Guedes - Bruni et al. 1997) e de 2.217 ind.ha-1 para outra área secundária na mesma região (Pessoa et al. 1997). Para a região de Ubatuba, SP, Silva & Leitão Filho (1982) obtiveram 640 indivíduos em 160 pontos amostrais, o que equivale a uma estimativa de 816 ind.ha-1. Outros estudos, desenvolvidos na Ilha Grande, RJ, apontam estimativas de 2.010 ind.ha-1 distribuídos, 18 famílias e 23 espécies (vegetação secundária com 03 anos de idade); e de 2.670 ind.ha-1, valor estimado à partir de 257 indivíduos amostrados, distribuídos em 17 famílias e 27 espécies (vegetação secundária com 03 anos) (Delamônica, P. 1997, comunicação pessoal).
7.1.2.3. Estrutura da Vegetação- topo
Os dados obtidos para o estudo da estrutura estão apresentados na Figura 6 e na Figura 7. Com relação à distribuição em classes de diâmetro para a Floresta 1, nota-se que 80% dos indivíduos estão concentrados nas classes até 25cm, enquanto que 6,2% dos indivíduos estão concentrados acima de 50cm de diâmetro. Para a Floresta 2 foi obtido 88% dos indivíduos nas classes até 25cm de diâmetro e 1,6% acima de 50cm. Para a Floresta 1 os maiores diâmetros encontrados ficaram com um exemplar de Ficus insipida com 111cm e outro de Hyeronima alchorneoides com 102cm. Para a Floresta 2 os maiores exemplares encontrados foram de Trichilia silvatica com 144cm e outro de Pourouma guianensis com 102cm.
Figura 6 - Distribuição das classes de diâmetro obtidas para os inventários realizados em Floresta 1 e Floresta 2.
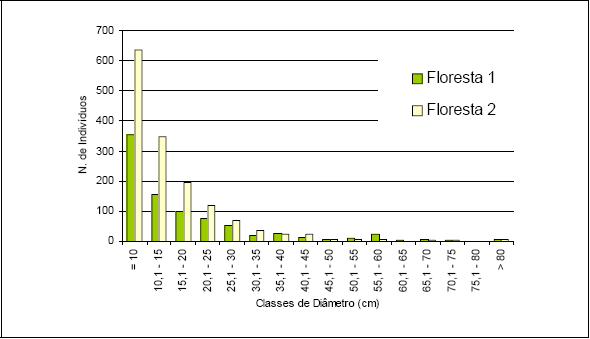
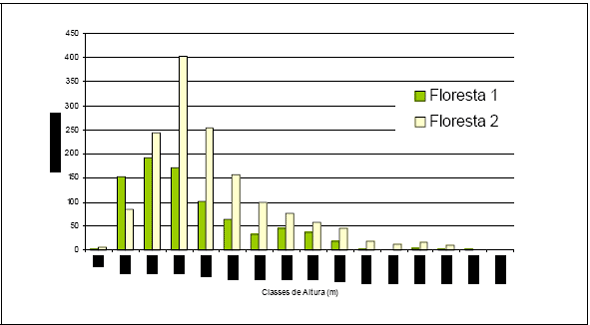
Figura 7 - Distribuição das classes de altura obtidas para os inventários realizados em Floresta 1 e Floresta 2.
Quintela & Louzada, 1993, encontraram na região áreas florestais em vários estágios de sucessão, em que a estrutura com distribuição à esquerda, mostra ser área mais jovem que a área com curva de distribuição à direita. Este fato também está evidenciado nas áreas amostradas, neste estudo.
Em relação à distribuição em classes de altura a Figura 7, resume os resultados obtidos, sendo que as duas florestas apresentam 3 estratos bem definidos. A Floresta 1 tem uma curva de distribuição de alturas mais à direita, evidenciando uma floresta em estágio de sucessão mais nova. A maioria dos indivíduos estão distribuídos nas classes mais baixas até 10 m de altura, distribuídos em 46 espécies, representando 45,5% do total de espécies ocorrentes. Somente 14,8% das espécies apresentam indivíduos com até 35 m, conforme pode ser observado na Tabela 6.
A Floresta 2, apresenta uma curva de distribuição de alturas mais à esquerda, mas também fica evidenciado que esta área florestal não é uma Floresta Primária, remanescente. Nesta floresta, a maioria dos indivíduos também ocorre nas duas primeiras classes até 10m de altura e até 20m, distribuídos em 38,6% e 36,1% do total de espécies, respectivamente. Destaca-se nesta área que 25,2% das espécies apresentam indivíduos com alturas até 35m, conforme pode ser observado na Tabela 7.
Assim, nota-se que as duas áreas florestais amostradas, são florestas em estágio de sucessão. A Floresta 1 pode ser classificada como uma capoeira e a Floresta 2, como uma floresta secundária recente. O mesmo fato é evidenciado pela curva de distribuição dos DAPs, apresentados na Figura 6.
Tabela 6 - Distribuição das famílias obtidas para a Floresta 1, de acordo com as alturas máximas obtidas neste levantamento.
Clique aqui para visualizar a tabela 6
Tabela 7 - Distribuição das famílias obtidas para a Floresta 2, de acordo com as alturas máximas obtidas neste levantamento.
Clique aqui para visualizar a tabela 7
A espécie Euterpe edulis (Arecaceae), desenvolve-se em florestas com melhor estado de conservação (ambientes sombreados), foi obtida com uma densidade de 49 ind. ha-1 e 54 ind. ha-1 para Floresta 1 e Floresta 2, respectivamente. Siqueira (1994, In: Guedes-Bruni et al. 1997) menciona que esta espécie aparece entre as cinco primeiras em cerca de 46% dos inventários realizados na costa brasileira. Na região conhecida como praia do Mamede, onde parte deste levantamento ocorreu, foram encontrados indícios da presença de palmiteiros. Não foram encontradas nas áreas amostradas espécies ameaçadas de extinção.
Na Tabela 8 estão listadas as espécies amostradas que têm uso econômico na região, além do nome específico, listou-se também o nome vulgar, geralmente utilizado na região para cada espécie.
Tabela 8 - Espécies de uso econômico
Clique aqui para visualizar a tabela 8
A região da Costa Verde apresenta uma das maiores áreas contínuas florestadas do estado e avaliando fotografias do entorno da CNAAA datadas da década de 70, pode-se constatar que a Floresta Ombrófila Densa, existente nas vertentes próximas, apresentou um aumento em sua extensão, e provavelmente avançaram na sucessão ecológica uma vez que a taxa de ocupação humana nesta região é baixíssima.
A formação considerada neste estudo foi a submontana (localizada entre 50 e 500m de altitude), com variações em seu estado de conservação. Esta variação observada em campo foi comprovada através dos resultados obtidos.
As áreas inventaridas possuem conjuntos de características que permitem individualizá-las tanto quanto a sua composição específica como estrutura da vegetação e estágio de sucessão. As duas florestas amostradas estão em estágio de sucessão, sendo que a Floresta 2 se encontra num estágio mais avançado, podendo-se dizer que já se encontra em fase de Floresta Secundária e que a Floresta 1 está em fase de capoeira avançada (Tabela 9).
Tabela 9 - Principais parâmetros sobre os estágios de sucessão das áreas avaliadas.
| Valores de vários parâmetros |
Floresta 1 |
Floresta 2 |
| Localização |
Junto à Estrada para Usina |
Fazendas antigas abandonadas |
| Número de Indivídios |
840 |
1.488 |
| Número de Famílias |
39 |
47 |
| Número de Gêneros |
70 |
101 |
| Número de Espécies |
101 |
205 |
| Principais Famílias |
Euphorbiaceae
Rubiaceae
Myrtaceae
Leguminosae
Melastomataceae
#1 Famílias encontradas em áreas mais jovens |
Myrtaceae
Rubiaceae
Lauraceae
Sapotaceae
Leguminosae |
| Índice de Diversidade |
1,77 |
2,03 |
| Percentual de espécies acima de 20m |
14,8% |
25,2% |
Para as vertentes próximas à CNAAA, onde temos a Floresta 1, a mata caracteriza-se por menores valores tanto para altura como diâmetro, sendo sua riqueza de espécies inferior à da Floresta 2. Já na bacia do córrego praia Brava, onde parte deste estudo se desenvolveu (Floresta 2), existem claros indícios da pretérita presença humana, porém ao mesmo tempo são observados espécimes de grande porte ou de espécies comumente observadas em florestas melhor preservadas, indicando uma extração seletiva e uma posterior recuperação, ou ainda pequenos mosaicos remanescentes da floresta anterior.
7.1.3. Restinga - topo
Variações do nível do mar, que ocorreram entre sete e dois mil anos atrás no período quaternário, no Holoceno, formaram ao longo do que hoje é a costa brasileira, vasta planície sedimentar arenosa (Suguio & Martin, 1990).
Em conseqüência do avanço e do recuo das águas do mar - transgressões e regressões marinhas - formaram-se planícies constituídas por uma sucessão de cordões arenosos. Após a deposição de areia, as tempestades, as correntes litorâneas, os ventos e as ondas passaram a modelar uma topografia complexa e diversificada (Araújo 1987; Suguio & Martin, 1987; Suguio et al. 1985). A esse conjunto de formações geomorfológicas e as comunidades biológicas que aí ocorrem, se dão o nome genérico de restinga (Rizzini, 1979).
As restingas na costa brasileira ocupam cerca de 79% de sua extensão total (Lacerda et al.,1993) sendo encontradas principalmente no litoral oriental e meridional.
O litoral brasileiro e a localização das planícies arenosas, segundo Araújo (1992), pode ser caracterizado, de acordo com a geomorfologia e o clima, como se segue (Figura 8).
Figura 8 - Regiões do litoral brasileiro, baseado na geomorfologia e clima (apud Araújo, 1992).
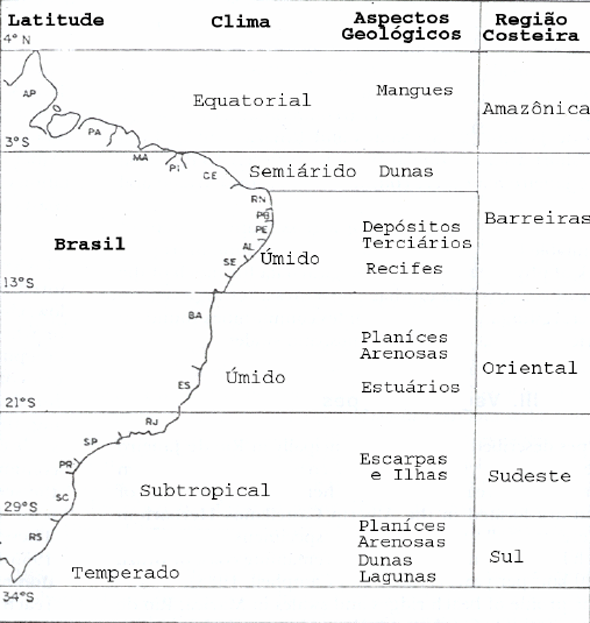
Na parte equatorial predominam os manguezais, principalmente por causa dos sedimentos trazidos pelo rio Amazonas e da amplitude das marés. Porém existem restingas, como por exemplo, na Ilha do Algodoal, na Baía de Marajó, no Pará.
No nordeste, no trecho de clima semi-árido, predominam os grandes campos de dunas. Ao sul do cabo São Roque, no Rio Grande do Norte, temos um clima úmido, com recifes, planícies arenosas estreitas, tabuleiros (formações terciárias) e costas escarpadas.
A partir do Recôncavo Baiano, no litoral sudeste, clima úmido, temos planícies costeiras bem desenvolvidas, principalmente no sul da Bahia, norte do Espírito Santo e Rio de Janeiro.
Na região subtropical existem muitas escarpas pré-cambrianas e ilhas, com uma linha costeira irregular.
No Sul, de clima temperado, ocorrem muitas restingas, campos de dunas e grandes sistemas de lagunas.
A diversidade geomorfológica, aliada ao gradiente ambiental do mar para a terra permite o estabelecimento de um notável mosaico de formações ambientais. Esta vegetação abrange uma diversidade de comunidades relacionadas a variações topográficas e microclimáticas e também a fatores temporais como história geológica e estágios de sucessão ecológica (Dillenburg et al.,1992).
Recobrem as planícies costeiras uma gama de comunidades com características próprias e diversificadas, tais como: vegetação rala de gramíneas, vegetação baixa agrupada em moitas, áreas de brejo com plantas aquáticas, matas fechadas. Todas abrangendo uma flora e fauna rica e variada, atualmente, restrita algumas vezes, a manchas remanescentes (Araújo, 2000).
Vários nomes de antigos logradouros referem-se à fauna das restingas, tais como Araruama (“viveiro de lontras”), Camorim (robalo), Catete (cateto, porco do mato), Guaratiba (“local dos guarás”), Jacarepaguá (“lagoa dos jacarés”), Jacarepiá (“pilha de jacaré”), Inhaúma (anhuma, ave), etc. Assim a restinga parece ser um ambiente bastante atraente para a fauna, devido a grande riqueza florística e elevada produção de flores e frutos durante todo o ano (Maciel, 1984).
O estado do Rio de Janeiro está situado em uma faixa tropical que representa a transição entre o litoral oriental dominado por falésias, recifes e deltas de grandes rios e o litoral sudeste dominado pelas serras que mergulham no mar, formando uma costa recortada e repleta de ilhas. Esta posição geográfica privilegiada proporciona para o Estado uma grande variedade de habitats, que por sua vez, dão origem a uma alta diversidade florística. Quatro grandes tipos de vegetação formam a cobertura vegetal do estado: a Floresta Atlântica, as Restingas, os Campos de Altitude e os Manguezais (Araújo & Maciel, 1998). As restingas do Estado ocupam uma área aproximada de 1200 km2, ou seja, 2,8 % do território do estado (Araújo & Maciel, op. cit.).
Nas baías do extremo oeste do estado do Rio de Janeiro, a retificação de trechos de costa por meios de restingas não é freqüente pela escassez de massas arenosas, a não ser na extremidade oriental, na zona de Sepetiba, onde os Rios Itaguaí e Guandu forneceram amplos volumes de areia, que em condições paleográficas privilegiadas, possibilitaram a formação da grande restinga da Marambaia. No restante desta costa, são raros os vales marítimos que abrigam planícies arenosas nas brechas montanhosas.
7.1.3.1. A Restinga na Região do Empreendimento - topo
Especificamente as baías na região de Angra dos Reis têm também como uma das características, a ausência de ondas, sendo mínimas as possibilidades de abrasão nessas águas protegidas pela Ilha Grande e que banham uma costa desfavorável à formação de restingas, onde predominam os pequenos rios torrenciais (Lamego, 1945). As planícies costeiras, com exceção da praia do Sul localizada na Ilha Grande, são geralmente, estreitas, caracterizadas por uma vegetação de praia, seguida por uma transição de restinga com floresta atlântica (Figura 9).
Figura 9 - Vista da praia da Batanguera onde foi realizada a presente caracterização. Área vizinha à Vila Residencial de Mambucaba (Aerofoto de 1987).

A comunidade de vegetação de praia é constituída de espécies principalmente herbáceas, comuns a todo o litoral brasileiro, como Alternanthera maritima, Ipomoea pes-caprae, Panicum racemosum, etc., que formam um emaranhado de rizomas e estolões sobre a areia.
Figura 10 - Vista do rio Mambucaba, entre a Vila Residencial de Mambucaba (à esquerda) e a propriedade onde está instalada a Pousada do Bosque. Em destaque a área onde foi realizada a presente caracterização (Aerofoto de 1987).

A restinga, propriamente dita, apresenta uma vegetação arbustiva - arbórea, sendo comumente encontrado Clusia sp, Cupania oblongifolia, Guarea macrophylla, Ouratea cuspidata, Schinus terebinthifolius, Tapirira guianensis, entre outras espécies características deste ecossistema. A altura dos indivíduos torna-se maior na área de transição com a mata de encosta. Nas áreas com impactos antrópicos instala-se, rapidamente, a embaúba(Cecropia sp.) como espécie característica.
Nas encostas rochosas e nas escarpas próximas ao mar, desenvolve-se uma vegetação com exemplares com características xeromórficas, como Cactaceae e Bromeliaceae, devido à intensa exposição solar, excesso de evaporação e pouca profundidade do solo. Certas palmeiras, como por exemplo, Attalea dubia, são comuns nestes costões.
Historicamente, após o abandono das grandes culturas na região os principais habitantes do litoral passaram a ser os caiçaras, que no passado foram indivíduos à margem da sociedade que se embrenharam pelos litorais, miscigenando-se com os remanescentes indígenas e posteriormente com os negros; adotando muitas práticas indígenas, como manejo agrícola, pescas de canoas no mar ou nas lagoas e a utilização dos produtos naturais das restingas, como alimentação, cestaria, remédios, tanoaria, tinturas (Lamego, 1974). A cultura destas populações era baseada na pesca e nas roças de subsistência (Oliveira & Coelho Neto, 1996; Oliveira, 1999).
Antes da Rodovia Rio-Santos, ou seja, até o início da década de setenta, o litoral sul fluminense e norte paulista estiveram praticamente isolados dos grandes centros urbanos, em conseqüência, a degradação dos ecossistemas não se encontrava em grande escala (UFRRJ/IEF/Pró-Natura 1993).
Atualmente, a vegetação litorânea está bastante alterada, em função do desmatamento, pela abertura de estradas com lançamento de aterro nas praias, por empreendimentos imobiliários e instalações industriais, além da crescente exploração turística. Deste modo é difícil sabermos as feições originais da vegetação do litoral de Angra dos Reis.
Do período áureo da cana e do café, restam ruínas de antigas fazendas, como da Fazenda Bracuí, onde se localizava um dos antigos cais, atual condomínio Porto Marina Bracuhy. Assim também no antigo porto de Mambucaba, está a Vila Histórica de Mambucaba e o Eco Resort Hotel do Bosque. Empreendimentos hoteleiros e centros urbanos periféricos se encontram em outras áreas da planície litorânea, como Frade, Trindade, Laranjeiras.
As restingas da região continental de Angra dos Reis, já bastante alteradas, são pouco conhecidas, sendo necessário levantamentos de dados históricos e atuais, para fornecer subsídios para futuros projetos de preservação e recuperação, principalmente visando o potencial turístico local.
A área de abrangência do diagnóstico da vegetação das restingas do entorno da Central Nuclear Almirante Álvaro Alberto - CNAAA, teve que ser expandida devido a uma combinação de fatores geomorfológicos e históricos desta região costeira.
O litoral sul do Estado do Rio de Janeiro é muito recortado, com numerosas enseadas e ilhas; sendo que as escarpas íngremes da Serra do Mar constituem na maioria das vezes, a própria linha da costa, impedindo o desenvolvimento das extensas planícies costeiras de restingas (Muehe & Valentini, 1998).
No raio de 5 km da Usina Nuclear de Angra dos Reis, as praias se caracterizam por formações de Mata Atlântica que recobrem em um contínuo a faixa litorânea, tais como a Prainha, ou por pequenas faixas de areia entre costões rochosos, por exemplo, a praia Vermelha. Para selecionar as áreas de restinga que melhor representassem este ecossistema na região, foram percorridas todas as áreas desde o rio Bracuí até Tarituba.
Algumas planícies arenosas mais amplas, como a formada pelo rio Bracuí, sofreram um processo de ocupação e conseqüente exploração da área, que remonta à época colonial (Lamego, 1945, 1974), não se encontrando atualmente vestígios das formações originais de restingas.
Na planície costeira formada pelo rio Mambucaba, na divisa entre os municípios de Angra e Parati, encontra-se uma formação geológica de depósitos arenosos, (Domingues et al., 1976), ainda com elementos que caracterizam uma vegetação de restinga.
No extremo desta praia, em direção ao sul, localiza-se a praia da Vila Residencial de Mambucaba, uma área antropizada. Porém adjacente a esta, a praia da Batanguera é formada por uma ampla planície arenosa, entre duas formações rochosas, também com elementos de vegetação de interesse para o trabalho em questão.
Para execução da caracterização das restingas nas áreas de influência, inicialmente foi percorrido o litoral, tomando-se como marco zero as Usinas de Angra 1 e 2, em Itaorna, BR-101- Km 132, para uma caracterização das áreas com vegetação de restinga.
Na direção norte, no sentido Parati-Angra dos Reis, encontram-se pequenas praias, a Praia de Mambucaba; o Saco Piraquara de Fora, nas quais não há formação de restinga. Também a mata atlântica chega próxima à linha da costa como na praia do Frade. Também estão presentes os manguezais.
A planície arenosa formada pelo rio Bracuí, uma formação geológica mais ampla, propícia à formação de restinga, vem sendo alterada historicamente desde o período das grandes plantações de cana e café. Atualmente existem instalados nesta área condomínios, porto e marina.
Na direção sul, no sentido Angra-Parati, na praia Brava localiza-se a vila residencial da Eletronuclear.
O Saco Piraquara de Dentro é uma praia entre costões, caracterizada por uma vegetação rasteira de praia, com a mata atlântica de encosta dos morros adjacentes, chegando bem próxima à praia. Existe camping e atividades de pescadores.
A praia Vermelha é uma faixa de areia avermelhada com pequena extensão, entre costões rochosos. Possue casas construídas bem próximo ao mar.
Na Prainha, a mata também se apresenta como um contínuo dos morros próximos e a praia com vegetação rasteira. Aqui existe uma área de camping em meio às árvores, onde o sub-bosque foi removido, provavelmente para desfrute do sombreamento. Ainda podem ser encontradas pequenas manchas de mata com epífitas, a exemplo de aráceas e bromélias.
A área denominada praia do Hotel do Bosque (Figura 10), está localizada a cerca de sete quilômetros das usinas de Angra 1 e Angra 2, adjacente à foz do rio Mambucaba; delimitada pela Vila Histórica de Mambucaba e no extremo oposto pela Vila Residencial de Mambucaba habitada por funcionários da Eletronuclear. O processo de formação desta planície arenosa está relacionado ao rio Mambucaba, o de maior vazão na região. O acesso à área também pode ser realizado por barco, atravessando o rio, meio utilizado para o transporte de funcionários, hóspedes e visitantes do Eco Resort Hotel do Bosque.
A praia da Batanguera (Figura 9) inicia-se após a Vila Residencial, o acesso principal é próximo ao rochedo com um mirante e instalações de antenas, sendo alvo de freqüente visitação. A planície, com elementos de vegetação de restinga, se estende por cerca de um quilômetro entre duas formações rochosas.
Seguem-se pequenas planícies entre formações rochosas, como a de Tarituba, já distrito de Parati.
Para as finalidades deste trabalho e considerando-se as características especificas da região em termos físicos e bióticos, foram consideradas as áreas da praia do Hotel do Bosque e da praia da Batanguera como de relevante interesse em relação ao ecossistema de restinga.
Foram realizadas idas ao campo em junho e julho, sendo realizadas coletas extensivas de material botânico nas duas áreas de restinga. Devido ao período de realização do trabalho, parte das amostras foi coletada estéril (desprovidas de material reprodutivo: flor e fruto).
Os indivíduos coletados foram preparados em laboratório (herborização), de acordo com o procedimento descrito em Mori et al. (1985), e identificados através da metodologia clássica de morfologia comparada, chaves de identificação, literatura especializada, comparação com exsicatas e posterior auxílio de especialistas.
Foi elaborada uma listagem das espécies de plantas encontradas, incluindo além das espécies de restinga, outras a exemplo daquelas consideradas ruderais, para um melhor diagnóstico do “status” das áreas.
Estimou-se a dimensão de cada área e analisaram-se os possíveis padrões de zonação da vegetação, a partir de linhas-base, longitudinais e perpendiculares ao oceano, com a utilização de uma trena de 50 m.
Foram levantadas referências bibliográficas, sobre a área do litoral de Angra dos Reis com ênfase em restingas. São citados artigos sobre áreas adjacentes, tendo em vista a carência de bibliografia sobre a área especifica.
Compõem a lista, trabalhos gerais sobre aspectos geomorfológicos, botânicos, faunísticos e ecológicos de restingas, que possam ser de interesse para o estudo.
Na orla da praia de ambas as áreas amostradas é comum a presença da amendoeira (Terminalia cattapa), um espécie exótica comum em diversas regiões, que apresenta rápido crescimento e dispersão.
A primeira comunidade vegetal (psamófila-reptante), em ambas as áreas, compreende a região próxima ao mar, coberta por espécies estoloníferas, rizomatosas e reptantes. As espécies representativas desta comunidade são comuns na faixa de todo o litoral do Rio de Janeiro, tais como Alternanthera littorales, Hydrocotyle bonariens, Ipomoea pes-capre, Marisicus pedunculatus, Panicum racemosum, Paspalum maritimum, Sporobolus virginicus.
A largura da comunidade variou de 6 a 10 m, na restinga da praia da Batanguera e cerca de 2m na do rio Mambucaba. Estando esta região ainda sobre influência dos efeitos das marés, as espécies ocorrentes são importantes na fixação da areia. A espécie que apresentou maior cobertura (75%) foi Ipomoea pes-capre. O sucesso desta espécie parece estar relacionado com sua forma de vida, uma herbácea que se desenvolve por crescimento clonal, bem adaptada a um substrato instável (Menezes, 1996).
Posterior a esta zona ocorre uma faixa arbustiva, na praia da Batanguera de largura de 7 a 25 m, constituída basicamente pela espécie Dalbergia ecastophylla e, algumas herbáceas e trepadeiras. Na restinga de Mambucaba, em uma faixa de características semelhantes, com cerca de 5 m, Senna pendula divide o espaço com a espécie arbustiva D. ecastophylla, ocorrendo em menor quantidade, na praia da Batanguera.
Na praia dos Coqueiros, a partir de uns 51 m da praia em direção à estrada, gramíneas daninhas (Brachiaria sp. Imperata brasiliensis, Panicum maximum) passam a infestar a área, tornando-a quase impenetrável por alcançarem até 1,5 m de altura.
Segundo moradores do local, há aproximadamente 20 anos atrás havia uma mata com muitas árvores, onde agora ocorrem as espécies invasoras supracitadas. Ainda existem remanescentes desta mata na parte sul, representados por manchas de espécies arbóreas de restinga, a exemplo de Andira legalis, Guapira opposita, Guarea macrophylla, Tapirira guianensis, Myrsine coriacea, Myrsine guianensis, Eugenia uniflora, Ouratea cuspidata, Psidium catteyanum; que ocorrem de resto como indivíduos isolados em meio ao capim. Esta faixa tem em média 150 m até a rodovia, sendo cortada por trilhas de acesso à praia.
Na restinga do rio Mambucaba, após a faixa arbustiva, encontramos uma mata de restinga, , com cerca de 50 m de largura. Caracteriza-se como um mata de restinga aberta, degradada, porém em um grau menor que na restinga anterior. A espécie Myrsine coriacea destaca-se nesta áreaocorrendo em agrupamentos, além de Schinus terebinthifolius, Erythroxylum aff. decidum e Cecropia pachystachya, esta última a embaúba, uma planta pioneira freqüente nas planícies litorâneas, inclusive muito utilizada na medicina popular.
A restinga da praia dos Coqueiros estende-se entre as duas formações rochosas por 1.100 m. A outra área de restinga, considerando-se o final da área urbanizada da Vila dos Funcionários, próximo ao quiosque do Hotel do Bosque como o seu inicio, compreende 480 m até a foz do rio Mambucaba.
Uma lista florística das duas áreas de estudo é apresentada na Tabela 10.
Tabela 10 - Lista de plantas vasculares das restingas de Mambucaba e da praia dos Coqueiros
Clique aqui para visualizar a tabela 10
As restingas, um dos ecossistemas associados da Mata Atlântica (Decreto Federal 750/93), estão protegidas na legislação sob o titulo genérico de Áreas de Formações Pioneiras, não incluindo somente sua flora e fauna, mas também o próprio solo.
Através da caracterização da vegetação realizada nas diferentes área visitadas não foi registrada a ocorrência de nenhuma espécie pertencente a qualquer uma das categorias da Lista Oficial de Flora Ameaçada de Extinção, Ibama - Portaria Nº 37-N, de 3 de abril de 1992.
Dentro da área deste estudo a praia dos Coqueiros é caracterizada por uma vegetação de restinga com transição para Floresta Atlântica, com muitas partes degradadas.
A praia do rio Mambucaba, também chamada de “praia do Hotel do Bosque”, é citada por Lamego (1945) como a principal área de formação restinga da região, também possuindo várias espécies invasoras em sua extensão.
A cobertura vegetal das restingas estudadas vem sendo alterada devido à utilização inadequada por parte dos visitantes, pela ocorrência de incêndios e posterior invasão de espécies exóticas ao ambiente.
Algumas das plantas remanescentes de restingas são caracterizadas pela rusticidade e pela capacidade pioneira, a exemplo das Myrsinaceae e Erythroxylaceae (Ribas, 1992), além da amplamente distribuída Schinus terenthifolius.
O número de espécies da família Compositae reflete as fisionomias abertas e a rápida ocupação por plantas oportunistas. Também foi significativo o grande número de espécies e indivíduos ruderais (e.g. Cayaponia martiana, Crotalaria pallida, Solanum argenteum, Spermacoce latifolia, Urena lobata) comuns em áreas degradadas. Na restinga de Mambucaba, registrou-se mesmo a presença de mamona (Ricinus communis).
Atualmente, em especial na praia dos Coqueiros, a visitação descontrolada está rapidamente degradando o ecossistema. Os incêndios, devido à proximidade da rodovia e descaso dos visitantes, também têm impedido a recuperação da restinga. Além da falta de controle da entrada de visitantes através da rodovia representar um risco mesmo à segurança das residências da Vila Residencial adjacente.
Apesar dos problemas supracitados as duas áreas destacam-se nas proximidades da área do empreendimento, por serem regiões litorâneas, com potencial para representar o ecossistema de restinga, propriamente dita, e restinga com transição para Floresta Atlântica. Podem ser ainda consideradas áreas remanescentes deste ecossistema, conservando parte da diversidade de plantas que provavelmente ocorria em tempos pretéritos. Além disto, soma-se aqui a importância do papel de corredores ecológicos para a Mata Atlântica.
7.1.4. Manguezal - topo
Este ecossistema é típico de áreas estuarinas tropicais e subtropicais, sendo regido por um sistema de variação de marés que o inunda duas vezes ao dia, onde a mistura das águas marinhas com as doces provenientes do continente profere um caráter salobro a estas águas.
A vegetação presente é halófita e desenvolve-se sobre um solo geralmente lamacento pobre em oxigênio, a qual forma um complexo Florestal Tropical Pluvial, caracterizando uma Vegetação com Influência Fluviomarinha dentre Áreas de Formação Pioneira segundo IBGE (1991), e como uma Vegetação Tropical Halofítica Latifoliada Perenifólia Sobre Vasa de Maré segundo Eiten (1983). Esta condição típica exerce um fator de seleção importante, quando referente ao tipo vegetal que colonizará este ambiente, ou seja, propicia que apenas algumas espécies sejam capazes de habitar este ecossistema.
Composto basicamente por uma vegetação de habitus arbustivo e/ou arbóreo, no Brasil, os manguezais estão representados por três gêneros ocorrentes em toda a sua costa onde o ecossistema se faz presente: Avicennia (mangue preto), Laguncularia (mangue branco) e Rhizophora (mangue vermelho), presentes em áreas onde o substrato é lamacento.
O ecossistema pode apresentar além do bosque, duas outras paisagens que caracterizam-no como área de relevante interesse ecológico - o apicum e o lavado. O apicum localiza-se nos terrenos de topografia mais elevada e não é banhado pelas marés diárias, onde pode ocorrer uma vegetação arbustiva e/ou arbórea dos taxa já citados, juntamente com Conocarpus (mangue de botão) e herbáceas como dos gêneros Paspalum (grama doce),Philoxerus, Salicornia, Spartina (praturá), Sesuvium (beldroega) e Sporobolus. À frente do bosque, uma outra paisagem do manguezal se faz presente, o lavado, uma faixa de lama descoberta durante a maré baixa que se estende até o limite da água, que às vezes pode apresentar o gênero Spartina.
Em muitos manguezais pode haver a presença de uma faixa de vegetação herbácea na face externa deste ecossistema, localizada na transição com o próximo ambiente, onde algumas espécies das famílias Amarilydaceae, Cyperaceae e Gramineae compõem o cenário, sendo que sua ocorrência depende dos fatores ambientais. Esta zona de mudança pode também ser composta por plantas arbustivas e arbóreas. Todos estes vegetais são considerados como espécies associadas, dentre os quais podem ser encontrados alguns representantes como: Acrostichum aureum e Acrostichum danaeifolium (samambaia do brejo), Cyperus spp., Dalbergia ecastophylla, Hibiscus pernambucensis (algodoeiro da praia), Schinus terebinthifolius (aroeira).
Associadas a vegetação lenhosa do manguezal, encontra-se exemplares epifíticos terrestres das Famílias Araceae, Bromeliaceae e Orchidaceae como Aechmea nudicaulis, Anthurium scandens, Cattleya sp., Solander sp., Tillandsia stricta, Tillandsia usneoides, Vriesea spp. e Oncidium spp., além da Divisão Pteridophyta. Uma outra comunidade relacionada com os mangues é o fitobenton, composto por uma flora ficológica macroscópica fixa aos troncos, raízes e pneumatóforos das árvores. Estas pertencem a duas Divisões, Chlorophycophyta e Rhodophycophyta, as quais são representadas por alguns gêneros como: Bostrychia, Catenella, Chaetomorpha, Cladophoropsis, Enteromorpha, Rhizoclonium e Ulvaria. Estas algas estão ligadas ao manguezal de modo direto, pois participam da cadeia alimentar da fauna aquática e da produção de oxigênio. podendo ser utilizadas como indicadores biológicos por participarem da produção de matéria orgânica deste ecossistema.
Os manguezais da região sul fluminense, outrora bem preservados por estarem localizados em um ambiente de solo lamacento, começaram a sofrer as primeiras modificações a partir da década de 70, época em que foi realizada a construção da BR-101, a qual separou bosques destes ecossistemas nos municípios da região sul-fluminense.
Praticamente, os municípios de Mangaratiba e principalmente de Angra dos Reis e Parati, sob o ponto de vista ambiental, podem ser divididos antes e após a existência desta rodovia. Outro grande fator de degradação foi o “Projeto Turis”, estudo encomendado a Embratur, que objetivava promover a ocupação ordenada preservando as belezas naturais da região. Porém na realidade, este projeto serviu como instrumento para a ocupação predatória da zona costeira do litoral sul fluminense, através do aterro de manguezais para a construção de marinas e loteamentos de luxo (Moscatelli, 2001).
Moscatelli et al.(1993) descreve que nos últimos 30 anos no município de Angra dos Reis, os manguezais foram reduzidos à 60% da área originalmente ocupada por este ambiente. Já os estudos realizados por Kjerfve & Lacerda (1993) apud NATRONTEC (op. cit.), relatam que aproximadamente 2.000 ha de manguezais já foram perdidos em toda a Baía de Ilha Grande, número que demonstra que mais de 50% das formações deste ecossistema sofreram os impactos da civilização.
Os manguezais situados no trecho entre Angra dos Reis e Tarituba representam 8,12% da área total do litoral sul do estado do Rio de Janeiro (Natrontec, op. cit.), e de um modo geral encontram-se antropizados, estando, portanto, descaracterizados quanto ao seu estado nativo.
A fitofisionomia de manguezal foi observada no município de Angra dos Reis, citando-se áreas como a do rio Mambucaba, Praia do Recife, Ilha do Jorge, rio Ariró, rio Bracuhy, cidade de Japuíba, Caiera, enseada de Porto Marisco e cidade de Angra dos Reis. A classificação destas paisagens no tocante a sua fisiografia é a do tipo de franja para os da Caiera, da cidade de Angra dos Reis, da enseada de Porto Marisco, da Praia do Recife e da Ilha do Jorge, tipo ribeirinho para os dos rios Ariró, Bracuhy e Mambucaba, e de bacia para o manguezal da cidade de Japuíba.
A vegetação arbustiva/arbórea deste trecho é composta por Avicennia schaueriana (mangue preto), Laguncularia racemosa (mangue branco) e Rhizophora mangle (mangue vermelho). Margeando estes vegetais, podem ser observados os taxa Acrostichum aureum (samambaia do brejo), Annona sp. (araticum), Hibiscus pernambucensis (guaxima), Philoxerus portulacoides, Schinus terebinthifolius (aroeira), Sesuvium portulacastrum e Typha domingensis (taboa).
A fitofisionomia de Manguezal também foi observada no município de Parati, localizada nas áreas do Saco, do rio Jorge Nunes e da praia da Jabaquara. Estes manguezais são classificados no tocante a sua fisiografia, como sendo do tipo franja. As condições ambientais destes Manguezais são médias, uma vez que existe a pressão de empresários para construírem sobre áreas deste ecossistema, além de outras já degradadas. A história de degradação deste ecossistema se iniciou, conforme mencionado anteriormente, a partir da construção da BR-101.
A vegetação das formações deste ecossistema em Parati apresenta característica arbustiva/arbórea, sendo composta por Avicennia schaueriana, Laguncularia racemosa e Rhizophora mangle. Margeando aos bosques desta comunidade vegetal, podem ser observados os taxa Acrostichum aureum, Annona sp., Hibiscus pernambucensis, Schinus terebinthifolius, e Typha domingensis. A Tabela 11 abaixo apresenta a listagem das espécies vegetais comumente observadas nos manguezais.
Tabela 11 - Listagem de espécies vegetais observadas nos manguezais dos municípios de Angra dos Reis e Parati.
| Família |
Espécie |
Nome Popular |
| Polypodiaceae |
Acrosticum aureum |
samambaia do brejo |
| Polypodiaceae |
Acrosticum danaeifolium |
samambaia do brejo |
| Bromeliaceae |
Aechmea nudicaulis var. nudicaulis |
gravatá ou bromélia |
| Araceae |
Anthurium scandens |
- |
| Annonaceae |
Annona sp. |
araticum |
| Avicenniaceae |
Avicennia schaueriana |
mangue preto |
| Orchidaceae |
Catasetum sp. |
orquídea |
| Orchidaceae |
Cattleya sp. |
orquídea |
| Cyperaceae |
Cladium jamaicense |
- |
| Cyperaceae |
Eleocharis sp. |
junco |
| Cyperaceae |
Fimbristylis spadicea |
- |
| Malvaceae |
Hibiscus pernambucensis |
algodoeiro da praia |
| Malvaceae |
Hibiscus tiliaceus |
algodoeiro da índia |
| Combretaceae |
Laguncularia racemosa |
Mangue branco |
| Leg. Papilionoideae |
Machaerium sp. |
- |
| Melastomataceae |
Miconia cf. hymenonervia |
- |
| Myrtaceae |
Myrcia multiflora var. glaucens |
- |
| Orchidaceae |
Oncidium spp. |
orquídea |
| Orchidaceae |
Pleurothallis spp. |
orquídea |
| Polygalaceae |
Polygala paniculata |
- |
| Orchidaceae |
Polystachya sp. |
orquídea |
| Anacardiaceae |
Schinus terebinthifolius |
aroeira |
| Cactaceae |
Rhipsalis grandiflora |
cactos macarrão |
| Rhizophoraceae |
Rhizophora mangle |
mangue vermelho ou sapateiro |
| Bromeliaceae |
Tillandsia stricta |
gravatá ou bromélia |
| Bromeliaceae |
Tillandsia usneoides |
barba de velho |
| Sheuchzeriaceae |
Triglochin sp. |
- |
| Bromeliaceae |
Vriesea sp. |
gravatá ou bromélia |
Fonte: NATRONTEC (1998).
No intuito de efetuar uma caracterização dos manguezais mais próximos a CNAAA foram selecionadas cinco localidades onde ocorrem estes ecossistemas.
São estas: o manguezal do Bracuí, o manguezal da praia do Recife (Figura 11), o manguezal da Ilha do Jorge Grego (Figura 11), da Pousada do Bosque (Figura 12), e do rio Ariró (Figura 13).
Foram realizadas no total 3 campanhas de campo distribuídas nos meses de maio (3 dias), outubro (4 dias) e novembro (2 dias).
Nos manguezais, citados anteriormente, foram selecionados trechos onde foram dispostos perfis amostrais perpendiculares à linha d’água. O comprimento de cada perfil foi diferenciado em função da extensão destes ecossistemas.
Em cada perfil, realizaram-se amostragens em parcelas com 10 m de lado com distância de 50 m entre cada uma delas.
Para cada espécime das parcelas foram coletados dados relativos a sua altura, circunferência à altura do peito (CAP), presença de indivíduos jovens, presença de indivíduos mortos e presença de alterações ambientais, conforme metodologia descrita por Schaeffer-Novelli & Cintron (1983).
Também foram realizadas observações sobre o tipo de sedimento e os representantes da fauna presentes nas parcelas. Os dados coletados em campo foram anotados em planilha e transportados para laboratório. Cada parcela foi posicionada com auxílio de GPS marca Garmin.
No laboratório, os dados coletados foram transferidos para o computador onde se calculou o Diâmetro à altura do peito (DAP), a área basal e a densidade.
Figura 11 - Localização das áreas onde foram feitos os levantamentos de campo: manguezal do Recife (Aerofoto de 1987).

12 - Localização das áreas onde foram feitos os levantamentos de campo: manguezal da Pousada do Bosque.

Figura 13 - Localização das áreas onde foram feitos os levantamentos de campo: manguezal do rio Ariró. Aerofoto de 1987.

As informações levantadas no campo permitiram descrever os perfis trabalhados da seguinte maneira:
- o manguezal do Bracuí foi caracterizado por 8 parcelas totalizando uma distância aproximada de 400 m. A espécie predominante foi Laguncularia racemosa (mangue branco). O sedimento apresentou características areno-lamosas;
- o manguezal da Praia do Recife foi estudado através do posicionamento de 7 parcelas amostrais, totalizando 400 m até atingir a região de transição. A espécie dominante foi Rhizophora mangle (mangue vermelho ou sapateiro). O substrato também apresentou características areno-lamosas;
- o manguezal da Ilha do Jorge foi avaliado pela disposição de 5 parcelas, onde percorreu-se 250 m. A espécie que dominou foi Rhizophora mangle (mangue vermelho ou sapateiro). O sedimento foi composto por areia e lama;
- o manguezal do Ariró foi estudado através do posicionamento de 6 parcelas amostrais, totalizando aproximadamente 330 m até atingir a região de transição. A espécie dominante foi Laguncularia racemosa (mangue branco). O substrato também apresentou características areno-lamosas;
- o manguezal da Pousada do Bosque foi avaliado pela disposição de 3 parcelas, onde percorreu-se 150 m. A espécie que dominou foi Laguncularia racemosa (mangue branco) e o sedimento local composto por areia e lama.
Os totais observados de cada espécie estão apresentados na Tabela 12. Os resultados obtidos demonstraram que, de um modo geral, os bosques estudados nos cinco perfis apresentaram predominância de Rhizophora mangle (mangue sapateiro) com 444 exemplares, e Laguncularia racemosa (mangue branco) com 453 exemplares enquanto somente 85 exemplares de Avicennia schaueriana (mangue preto) foram observados, concentrados no manguezal do rio Ariró.
Tabela 12 - Total de espécimes encontrados nas parcelas amostrais.
| >Perfis |
Rhizophora mangle |
Laguncularia racemosa |
Avicennia schaueriana |
Número total
de indivíduos |
| Bracuí |
66 |
123 |
0 |
189 |
| Praia do Recife |
223 |
69 |
16 |
308 |
| Ilha do Jorge |
121 |
25 |
17 |
163 |
| Ariró |
44 |
121 |
52 |
217 |
| Pousada do Bosque |
0 |
115 |
0 |
115 |
| Total |
454 |
453 |
85 |
992 |
O posicionamento geográfico das Parcelas de cada perfil está apresentado na Tabela 13.
Tabela 13 - Coordenadas geográficas das parcelas estudadas nos manguezais situados na AID.
Bracuí |
| Parcela 1 |
S 22º 57’ 17, 2” |
W 44º 22’ 55,7” |
| Parcela 2 |
S 22º 57’15,6” |
W 44º 22’ 55,5” |
| Parcela 3 |
S 22º 57’13,5” |
W 44º 22’ 55,1” |
| Parcela 4 |
S 22º 57’6” |
W 44º 22’54,7” |
| Parcela 5 |
S 22º 57’9,7” |
W 44º 22’ 55,1” |
| Parcela 6 |
S 22º 57”8,1” |
W 44º 22” 55,7” |
| Parcela 7 |
S 22º 57’6,6” |
W 44º 22’ 57,3” |
| Parcela 8 |
S 22º 57’5” |
W 44º 22’ 58,7” |
Praia do Recife |
| Parcela 1 |
S 22º 56’ 42” |
W 44º 25’ 3,3” |
| Parcela 2 |
S 22º 56’ 41,2” |
W 44º 25’ 5,3” |
| Parcela 3 |
S 22º 56’ 56,0” |
W 44º 25’ 3,3” |
| Parcela 4 |
S 22º 56’ 38,4” |
W 44º 25’ 8,3” |
| Parcela 5 |
S 22º 56’ 36,8” |
W 44º 24’ 51,1” |
| Parcela 6 |
S 22º 56’ 34,4” |
W 44º 24’ 52,1” |
| Parcela 7 |
S 22º 56’ 33” |
W 44º 24’ 52,2” |
Ilha do Jorge |
| Parcela 1 |
S 22º 56’ 37,7” |
W 44º 24’ 51,8” |
| Parcela 2 |
S 22º 56’ 38,8” |
W 44º 24’ 32,1” |
| Parcela 3 |
S 22º 56’40,0” |
W 44º 24’ 30,1” |
| Parcela 4 |
S 22º 56’41,0” |
W 44º 24’ 27,8” |
| Parcela 5 |
S 22º 56’ 42,2” |
W 44º 24’ 24,7” |
Ariró |
| Parcela 1 |
S 22º 55’24,9” |
W 44º 20’ 33,9” |
| Parcela 2 |
S 22º 55’22,4” |
W 44º 20’ 34,8” |
| Parcela 3 |
S 22º 55’ 20,5” |
W 44º 20’ 35,7” |
| Parcela 4 |
S 22º 55’ 18,6” |
W 44º 20’ 36,1” |
| Parcela 5 |
S 22º 55’ 16,5” |
W 44º 20’ 36,3” |
| Parcela 6 |
S 22º 55’ 14,4” |
W 44º 20’ 36,5” |
Pousada do Bosque |
| Parcela 1 |
S 23º 01’ 25,1” |
W 44º 31’ 32,2” |
| Parcela 2 |
S 23º 01’ 21,7” |
W 44º 31’ 36,5” |
| Parcela 3 |
S 23º 01’ 19,2” |
W 44º 31’ 36,4” |
7.1.4.1. Manguezal do Bracuí - topo
Distribui-se ao longo de toda a praia de Bracuí acompanhando a influência das águas do Rio Bracuí. Este manguezal, que no passado possuía uma distribuição mais ampla, foi reduzido paulatinamente a partir da década de 80 sofrendo um acelerado processo de degradação, causado pela construção de condomínios, de marinas, de residências e de corte de madeira.
Percorrendo o manguezal é possível observar facilmente os reflexos oriundos da ação antrópica. Um dos resultados deste processo é a ocupação de outras espécies vegetais nos locais de ocorrência dos mangues.
O lado direito da praia de Bracuí encontra-se totalmente antropizado com presença de residências construídas sobre áreas de manguezal e vazamento de lixo. Apesar destes fatos pode-se observar um bosque de Avicennia schaueriana (mangue preto) bem desenvolvido, instalado sobre a areia e localizado na porção frontal da praia. Seu desenvolvimento foi favorecido certamente pela existência de um riacho que drena águas doces neste local.
Percorrendo a praia pode-se observar que a área de manguezal estabelece em diversos trechos contato com a areia com ocorrência de espécies invasoras e típicas de restinga. A exceção é o trecho onde desemboca rio localizado aproximadamente na parte central da praia. No lado esquerdo da praia o manguezal do Bracuí se encontra com o manguezal da Enseada dos Girassóis. Este último foi completamente degradado no final da década de 80 pelo empreendimento chamado Porto Abrigo dos Reis, que previa a construção de um condomínio e uma marina. Esta área foi dragada em alguns trechos e aterrada em outros. Boa parte da vegetação que secou foi vítima de incêndio e corte de madeira. Apesar do embargo da obra nenhuma medida de recuperação foi implementada nos manguezais da Enseada dos Girassóis que ao longo dos anos se recuperaram naturalmente.
O ponto selecionado para o posicionamento das parcelas foi localizado aproximadamente na porção central da Praia do Bracuí. Foram efetuadas observações em 8 parcelas localizadas conforme as coordenadas apresentadas na Tabela 13.
A parcela 1 apresentou exclusivamente exemplares de Laguncularia racemosa onde mensurou-se a ocorrência de 37 indivíduos. A altura dos exemplares variou entre 2,7 m e 4,7 m com média de 3,88 m (Tabela 14).
A classe diamétrica que mais contribuiu para a área basal desta parcela foi o DAP ³ 2,5 e < 5 com resultado de 0,03 m2 /0,01ha, que indicou a dominância de exemplares jovens (Tabela 14).
Tabela 14 - Número de indivíduos e valores de altura obtidos nas amostragens realizadas no perfil do Manguezal do Bracuí.
Parcela |
N |
Altura do bosque (m) |
| Superior |
Inferior |
Média |
| 1 |
37 |
4,7 |
2,7 |
3,88 |
| 4 |
32 |
7,7 |
2,6 |
5,44 |
| 5 |
26 |
9,8 |
3,8 |
6,69 |
| 6 |
41 |
8,2 |
2,8 |
5,13 |
| 7 |
21 |
6,7 |
3,1 |
4,93 |
| 8 |
32 |
5,5 |
2,8 |
4,01 |
Pode-se observar um elevado número de exemplares mortos nesta parcela em função do corte de árvores para aproveitamento a madeira.
As parcelas 2 e 3 não foram amostradas em função do tamanho reduzido dos exemplares vegetais.
A altura das plantas observadas variou entre 70 cm e 2,0 m, tamanhos inferiores ao definido na metodologia do estudo, fato que justifica não terem sido estudadas. A espécie dominante nestas duas parcelas foi Laguncularia racemosa representada por exemplares jovens. Estas parcelas estão em estágio de recuperação em função do intenso processo de corte a que foi submetida a cobertura vegetal neste local. O corte da vegetação assemelha-se ao padrão observado nos manguezais da Baía de Guanabara, onde a vegetação da porção externa é mantida intacta, enquanto as porções internas são desmatadas, dificultando assim sua identificação para os órgãos de fiscalização.
Na parcela 4, observou-se a ocorrência de Laguncularia racemosa e Rhizophora mangle com dominância para a primeira espécie. A altura dos indivíduos desta parcela variou entre 2,6 m e 7,7 m com média de 5,44 m.
O DAP médio obtido foi de 4,17 cm. A maior contribuição de área basal foi da espécie Laguncularia racemosa. Com relação a classe diamétrica a maior contribuição foi do DAP ³ 2,5 e < 5 com 0,04 m2/0,01 ha, conforme demonstrado na Tabela 15.
A parcela 5 apresentou a ocorrência de Rhizophora mangle e Laguncularia racemosa onde foram mensurados 26 exemplares. A altura média das plantas foi de 6,69 m e variou entre 3,8 m e 9,8 m.
O DAP médio observado foi de 8,75 cm. A espécie que mais contribui para a área basal foi Rhizophora mangle e a classe diamétrica de maior contribuição para área basal foi do DAP ³ 10 com 0,09 m2/0,01 ha (Tabela 15).
Na parcela 6 observou-se a ocorrência de Laguncularia racemosa e Rhizophora mangle com a presença de 41 exemplares. A altura variou entre o valor mínimo de 2,8 m e máximo de 8,2 m, com média de 5,13 m (Tabela 14).
Observou-se nesta parcela uma grande mortandade de troncos de Laguncularia racemosa que representou 22 % do total de troncos estudados.
A espécie que mais contribuiu para a área basal foi Laguncularia racemosa. O DAP médio foi de 5,75 cm. A classe diamétrica que mais contribuiu para a área basal foi do DAP ³ 5 e < 10 com 0,123039 m2/0,01 ha (Tabela 14).
A parcela 7 foi composta por Rhizophora mangle (espécie dominante) e Laguncularia racemosa, onde foram amostrados 21 exemplares e observados 4 exemplares mortos. A altura média registrada foi de 4,93 m. O valor mínimo de altura foi de 3,1 m e o máximo de 6,7 m, conforme apresentado na Tabela 14.
Tabela 15 - Dados de estrutura relacionada a área basal no perfil do Bracuí.
P |
N.ind |
DAP
(médio) |
Rm |
Lr |
As |
Total |
< 2,5 |
>³ 2,5 e < 5 |
³ 5 e < 10 |
³ 10 |
| 1 |
37 |
3,18 |
0 |
0,052042 |
0 |
0,052042 |
0,005743 |
0,031033 |
0,015266 |
- |
| 4 |
32 |
4,17 |
0,003993 |
0,075577 |
0 |
0,079570 |
0,001808 |
0,041547 |
0,036216 |
- |
| 5 |
26 |
8,75 |
0,130499 |
0,043699 |
0 |
0,174198 |
- |
0,005007 |
0,070758 |
0,098434 |
| 6 |
41 |
5,75 |
0,090745 |
0,094941 |
0 |
0,185687 |
0,002062 |
0,026160 |
0,123039 |
0,034424 |
| 7 |
21 |
6,7 |
0,068573 |
0,025641 |
0 |
0,094214 |
- |
0,005040 |
0,054187 |
0,034986 |
| 8 |
32 |
6,82 |
0,103361 |
0,037845 |
0 |
0,141207 |
- |
0,010391 |
0,082617 |
0,048198 |
| Total |
189 |
- |
0,397171 |
0,277705 |
0 |
0,674875 |
0,003870 |
0,088144 |
0,366817 |
0,216044 |
OBS.: P - Parcela; Rm - Rhizophora mangle; Lr - Laguncularia racemosa; As - Avicennia schaeuriana
A maior contribuição para área basal foi da espécie Rhizophora mangle. A classe diamétrica de maior contribuição para área basal foi a do DAP ³ 5 e < 10 com 0,054187 m2/0,01 ha. O DAP médio desta parcela foi de 6,7 cm conforme demonstrado na Tabela 15.
A composição específica da parcela 8 foi representada por Rhizophora mangle e Laguncularia racemosa, com predominância da primeira espécie. Nesta parcela foram amostrados 32 exemplares, sendo observados apenas 2 mortos. A altura dos indivíduos variou entre 2,8 m e 5,5 m e a média foi de 4,01 m (Tabela 14).
Nesta parcela, a exemplo do observado na parcela 6, foi também constatada a mortandade de troncos de Laguncularia racemosa.
Rhizophora mangle foi a espécie que mais contribuiu para a área basal desta parcela. O DAP médio foi de 6,82 cm e a classe diamétrica que mais contribuiu para a área basal foi a do DAP ³ 5 e < 10 com 0,082617 m2/0,01 ha.
A partir dos dados de número de indivíduos apresentados pôde-se elaborar o gráfico apresentado na Figura 14, que demonstra a elevada contribuição da espécie Laguncularia racemosa ao longo das parcelas amostrais.
Figura 14 - Composição quantitativa das espécies nas parcelas amostrais do manguezal do Bracuí.
A análise dos dados da Tabela 15 revelou que maior contribuição para área basal total deste perfil foi a da classe diamétrica ³ 5 e < 10 com total de 0,366817 m2/0,01 ha, que indicou um ambiente em estado de regeneração avançada.
Com relação à fauna foi observada na vasa lodosa da Praia do Bracuí a atividade de mariscagem. A captura de mariscos nesta região é feita principalmente sobre Anomalocardia brasiliana (berbigão) e Tagelus plebeius (unha-de-velho). Também se observou inúmeros exemplares de Neritina virginica pastando sobre as algas.
No manguezal foi registrada nas parcelas amostrais a ocorrência dos gastrópodes Littorina (Scabra) angulifera (buso do mangue) e Melampus coffeus (cafezinho). Nos canais também foi observada a captura de exemplares de Lucina pectinata (ameixa).
Entre os crustáceos foram observadas as seguintes espécies: Uca (Minuca) rapax (chama-maré), Uca (Celuca) leptodactyla (chama-maré), Aratus pisonii (marinheiro), Goniopsis cruentata (aratu vermelho e preto), Chasmagnatus granulata (catanhém), Ucides cordatus (caranguejo uçá) e Cardisoma guanhumi (guaiamum). Ao longo das parcelas foram observadas inúmeras armadilhas para captura de Ucides cordatus (caranguejo uçá), conhecidas como laço, que indicam uma intensa atividade de pesca deste animal.
7.1.4.2. Manguezal da Praia do Recife - topo
Localiza-se próximo ao rio do Saco. Este manguezal é caracterizado por uma bela paisagem sendo muito procurado para atividades de lazer, como por exemplo, passeios de barco e banho de mar.
O acesso a este manguezal é feito através de uma estrada de terra que se comunica com a rodovia Rio - Santos. O trecho inicial desta estrada se localiza sobre uma área de mata secundária que está, aos poucos, sendo totalmente degradada. Ano após ano a mata está desaparecendo. Durante o trabalho expedito para elaboração deste relatório foi observado o corte de uma área de mata e drenagem de uma área brejosa no lado direito da estrada no sentido da praia. Avançando um pouco mais pela estrada pode-se constatar que a mesma foi construída sobre o manguezal, provocando modificações nas feições deste ecossistema. Os manguezais localizados ao lado da estrada não morreram, porque a mesma permite em momentos de maré cheia, a passagem de água por tubulões ou por cima da própria estrada, favorecendo a circulação das águas.
A comunidade da praia do Recife é formada basicamente por pescadores com aproximadamente um total de 30 famílias. A região também é freqüentada aos fins de semana e durante as férias escolares por pessoas que residem no Frade. Os pescadores sobrevivem basicamente de peixe, principalmente através da captura de tainha e corvina. Porém ocorre em pequena escala a captura de mariscos em frente a praia e/ou no manguezal, que são utilizados como complemento alimentar ou vendidos ocasionalmente. No passado a captura de mariscos era mais intensa em virtude da exploração e comercialização destes animais em grande escala para o Frade, Bracuí, Japuíba e Angra dos Reis. O principal molusco capturado era Anomalocardia brasiliana (berbigão). Atualmente a captura deste animal foi reduzida em virtude da mudança de residência da família que comercializava os mariscos.
O manguezal da Praia do Recife apresenta-se distribuído sobre um sedimento arenoso. Ao longo da Praia é possível observar diversos trechos onde este ambiente se desenvolveu com a presença de exemplares de até 5 m de altura, compostos por Avicennia schaueriana e Laguncularia racemosa.
Apesar da exuberância do Manguezal da Praia do Recife pode-se observar em diversos trechos o corte de madeira de mangue. A degradação do manguezal é mais intensa nos trechos localizados próximos às residências. Segundo informações obtidas com os pescadores locais a região está sendo objeto de especulação imobiliária, pois um empresário da região está comprando casas e terrenos com objetivo de transformar a praia em um condomínio fechado. A conseqüência deste processo se reflete na vegetação manguezal que está sendo cortada em diversos pontos.
O perfil amostral foi posicionado ao final do cordão arenoso no lado esquerdo da praia, localizado aproximadamente a uns 150 m antes da foz do Rio do Saco. As coordenadas geográficas das parcelas estão apresentadas na Tabela 13.
A parcela 1 foi caracterizada através da amostragem de 31 exemplares, onde observou-se a ocorrência de Rhizophora mangle, Laguncularia racemosa e Avicennia schaueriana. Na porção posterior, entre a parcela 1 e 2, foi observado um exemplar bem antigo de Avicennia schaueriana com 14 m de altura e 122 cm de CAP, valores bem superiores aos observados na parcela 1. A área ao lado da parcela apresentou diversos indivíduos mortos por desmatamento. A altura variou entre 2,6 m e 12 m com média de 5,65 m (Tabela 16).
Tabela 16 - Numero de indivíduos e valores de altura obtidos nas amostragens realizadas no perfil do Manguezal da Praia do Recife.
Parcela |
N |
Altura do bosque (m) |
| Superior |
Inferior |
Média |
| 1 |
31 |
12 |
2,6 |
5,65 |
| 2 |
35 |
11,2 |
3,2 |
6,35 |
| 3 |
36 |
5,5 |
2,6 |
3,38 |
| 4 |
91 |
7,6 |
2,5 |
4,96 |
| 5 |
40 |
10,5 |
2,5 |
5,85 |
| 6 |
53 |
11 |
2,5 |
4,64 |
| 7 |
22 |
7,5 |
2,8 |
5,33 |
A espécie de maior contribuição para área basal nesta parcela foi Rhizophora mangle (Tabela 16). O DAP médio obtido foi de 7,13 cm e a classe diamétrica que mais contribuiu para a área basal foi a do DAP ³ 10 com 0,168995 m2/0,01 ha.
Na parcela 2 a composição específica foi representada por Rhizophora mangle, Laguncularia racemosa e Avicennia schaueriana, totalizando 35 exemplares amostrados conforme demonstrado na Tabela 16. Dentro da parcela foram registrados 4 indivíduos de Rhizophora mangle mortos através de corte do tronco. Observou-se também próximo a parcela alguns exemplares de Hibiscus pernambucensis (algodoeiro da praia). A altura média do bosque foi de 6,35 m, com valor mínimo de 3,2 m e máximo de 11,2 m.
O DAP médio mensurado foi de 7,12 cm, bem semelhante ao DAP médio obtido na parcela 1. A classe diamétrica de maior contribuição para área basal foi a do DAP ³ 10 com 0,204590 m2/0,01 ha. A espécie que mais contribuiu com a área basal nesta parcela foi Rhizophora mangle (Tabela 17).
A parcela 3 foi representada por 36 exemplares das espécies Rhizophora mangle (dominante) e Laguncularia racemosa. Os valores registrados para altura do bosque variaram entre 2,6 m e 5,5 m. A altura média obtida foi de 3,38 m (Tabela 16).
A espécie de maior contribuição para área basal foi Rhizophora mangle. Com relação a classe diamétrica, a maior contribuição para área basal foi a do DAP ³ 5 e < 10 com 0,032436 m2/0,01 ha. O DAP médio registrado nesta parcela foi de 4,04 cm (Tabela 17).
Na parcela 4 foi registrada a ocorrência de Rhizophora mangle, Laguncularia racemosa e Avicennia schaueriana. O número de indivíduos amostrados foi o maior de todo o perfil com um total de 91 exemplares (Tabela 16). A altura média do bosque desta parcela foi 4,96 m, sendo que os valores de altura oscilaram entre 2,5 m e 7,6 m.
O DAP médio obtido foi de 4,13 cm, valor bem semelhante ao observado na parcela 3 (Tabela 17). A classe diamétrica de maior contribuição para área basal foi a do DAP ³ 2,5 e < 5 com 0,065866 m2/0,01 ha. A espécie Rhizophora mangle foi a que mais contribuiu para a área basal nesta parcela.
Tabela 17 - Dados de estrutura relacionados a área basal no perfil da Praia do Recife.
A parcela 5 apresentou a ocorrência de exemplares de Rhizophora mangle (mangue vermelho), Laguncularia racemosa e Avicennia schaueriana (mangue preto) onde mensurou-se a ocorrência de 40 indivíduos. A altura dos exemplares variou entre 2,5 m e 10,5 m com média de 5,85 m (Tabela 16).
A classe diamétrica que mais contribuiu para a área basal desta parcela foi o DAP ³ 10 com resultado de 0,273174 m2 /0,01ha, que indicou a dominância de exemplares maduros (Tabela 17). O DAP médio foi de 7,73 cm. Nesta parcela a espécie que mais contribuiu para a área basal foi Rhizophora mangle.
Na parcela 6 observou-se a ocorrência de Rhizophora mangle (mangue vermelho), Laguncularia racemosa e Avicennia schaueriana (mangue preto) com dominância para a primeira espécie. A altura dos indivíduos desta parcela variou entre 2,5 m e 11 m com média de 4,64 m (Tabela 16). O DAP médio obtido foi de 6,64 cm. A maior contribuição de área basal foi da espécie Rhizophora mangle. Com relação à classe diamétrica a maior contribuição foi do DAP ³ 5 e < 10 com 0,124452 m2/0,01 ha, conforme demonstrado na Tabela 17.
A parcela 7 apresentou a ocorrência de Rhizophora mangle e Laguncularia racemosa onde foram mensurados 22 exemplares. A altura média das plantas foi de 5,33 m e variou entre 2,8 m e 7,5 m (Tabela 16). O DAP médio observado foi de 7,86 cm. A espécie que mais contribui para a área basal foi Rhizophora mangle e a classe diamétrica de maior contribuição para área basal foi do DAP ³ 5 e < 10 com 0,082432 m2/0,01 ha (Tabela 17).
A Figura 15 apresenta a contribuição quantitativa de cada espécie ao longo do perfil, onde destaca-se a dominância de Rhizophora mangle em todas as parcelas.
Figura 15 - Composição quantitativa das espécies nas parcelas amostrais do manguezal da Praia do Recife.
Os valores de área basal, demonstrados na Tabela 17, indicaram que a maior contribuição para área basal total foi do DAP ³ 10 com 0,402145 m2/0,01 ha, fato que permitiu caracterizar este perfil como mais maduro estruturalmente.
A fauna observada no Manguezal da Praia do Recife foi representada por moluscos e crustáceos. Entre os moluscos foi observada a ocorrência de Neritina virginica, Littorina (Scabra) angulifera (buso do mangue), Tagelus plebeius (unha de velho), Anomalocardia brasiliana (berbigão), Lucina pectinata (ameixa), Iphigenia brasiliensis (tarioba), Teredo spp. (broca da madeira) e Ostrea rhizophorae (ostra do mangue).
Os crustáceos foram representados pelas seguintes espécies: Chthamalus spp. (craca), Balanus spp. (craca), Callinectes spp. (siri), Goniopsis cruentata (aratu vermelho e preto), Aratus pisonii (marinheiro), Uca spp. (chama-maré), Cardisoma guanhumi (guaiamum) e Ucides cordatus (caranguejo uçá). Nas parcelas amostrais foi observada uma grande quantidade de armadilhas para captura de Ucides cordatus conhecidas como laços, que constitui um forte indicativo de intensa pesca deste crustáceo na região. Ao percorrer outros trechos deste manguezal foi encontrado um acampamento, que segundo os pescadores locais, foi montado por caranguejeiros do Rio de Janeiro, que periodicamente visitam o Manguezal da Praia do Recife para capturar o caranguejo uçá.
7.1.4.3. Manguezal da Ilha do Jorge - topo
Localiza-se numa área contígua ao manguezal da praia do Recife. O acesso a este manguezal somente foi possível com apoio de um barco gentilmente cedido pelos pescadores da Praia do Recife. O perfil amostral do manguezal da ilha do Jorge foi posicionado ao lado de um canal de maré. As coordenadas geográficas de cada parcela estão apresentadas na Tabela 13.
Na parcela 1, a composição específica do bosque foi representada por Rhizophora mangle, Laguncularia racemosa e Avicennia schaueriana. A espécie dominante nesta parcela foi Rhizophora mangle.
Observou-se a presença de raízes de estresse em um indivíduo de Avicennia schaueriana. O número total de indivíduos amostrados foi de 49 (Tabela 18). A altura média do bosque foi de 3,97 m, sendo que os exemplares apresentaram a altura mínima de 2,6 m e a máxima de 12,5 m.
Tabela 18 - Número de indivíduos e valores de altura obtidos nas amostragens realizadas no perfil do Manguezal da Ilha do Jorge.
Parcela |
N |
Altura do bosque (m) |
| Superior |
Inferior |
Média |
| 1 |
49 |
12,5 |
2,6 |
3,97 |
| 2 |
21 |
11 |
5 |
9,46 |
| 3 |
20 |
10,2 |
2,9 |
6,29 |
| 4 |
30 |
9,5 |
2,8 |
6,53 |
| 5 |
43 |
7,5 |
2,6 |
5,40 |
A espécie de maior contribuição para área basal da parcela 1 foi Rhizophora mangle (Tabela 19). A classe diamétrica que mais contribuiu para a área basal foi do DAP ³ 10 com 0,054057 m2/0,01 ha. O DAP médio registrado de 3,7 cm.
Na parcela 2, a altura do bosque variou entre 5 e 11 m. O valor médio registrado para a altura dos exemplares foi de 9,46 m (Tabela 18). Foi observada a ocorrência de Rhizophora mangle e Avicennia schaueriana. O DAP médio obtido nesta parcela foi de 11,14 cm conforme apresentado na Tabela 19. Rhizophora mangle foi a espécie que mais contribuiu para a área basal. A classe diamétrica de DAP ³ 10 com 0,198814 m2/0,01 ha foi que mais contribuiu para a área basal.
Tabela 19 - Dados de estrutura relacionada a área basal no perfil da Ilha do Jorge
P |
N.
ind |
DAP
(médio) |
Rm |
Lr |
As |
Total |
< 2,5 |
> 2,5 e < 5 |
> 5 e < 10 |
> 10 |
1 |
49 |
3,7 |
0,043286 |
0,001904 |
0,071874 |
0,117064 |
0,008275 |
0,012791 |
0,041941 |
0,054057 |
2 |
21 |
11,14 |
0,210617 |
0 |
0,029220 |
0,239837 |
0 |
0,001146 |
0,039877 |
0,198814 |
3 |
20 |
10,31 |
0,135582 |
0,020547 |
0,061600 |
0,217729 |
0,000390 |
0,003398 |
0,028284 |
0,185658 |
4 |
30 |
8,46 |
0,129592 |
0,064016 |
0,019894 |
0,213501 |
0,000390 |
0,004291 |
0,071399 |
0,137421 |
5 |
43 |
6,68 |
0,127919 |
0,043150 |
0,004800 |
0,175870 |
0,000390 |
0,013797 |
0,124242 |
0,037441 |
Total |
163 |
|
0,646995 |
0,129617 |
0,187389 |
0,964001 |
0,009445 |
0,035423 |
0,305742 |
0,613390 |
O bosque da parcela 3 foi composto por Rhizophora mangle, Laguncularia racemosa e Avicennia schaueriana. O número de exemplares amostrados foi semelhante ao valor de indivíduos estudados na parcela 2. A altura do bosque variou entre 2,9 m e 10,5 m. O valor médio registrado para altura foi de 6,29 m (Tabela 18). O DAP médio obtido foi de 10,31 cm. A espécie de maior contribuição para a área basal foi Rhizophora mangle (Tabela 19). A classe diamétrica de maior contribuição foi a do DAP ³ 10 com 0,185658 m2/0,01 ha.
As espécies Rhizophora mangle, Laguncularia racemosa e Avicennia schaueriana foram registradas na parcela 4. A altura média dos 30 exemplares amostrados foi de 6,53 m. O valor mínimo mensurado para altura foi de 2,8 e o valor máximo foi de 9,5 m (Tabela 18). O DAP ³ 10 foi o que apresentou maior contribuição para área basal onde se registrou 0,137421 m2/0,01 ha. O DAP médio registrado foi de 8,46 cm. A maior contribuição para área basal foi registrada para a espécie Rhizophora mangle (Tabela 19).
Entre as parcelas 4 e 5, o bosque do manguezal da ilha do Jorge sofreu uma redução de altura e um aumento da densidade de exemplares, fatores característicos de um manguezal em regeneração, o que dificultou o acesso até o ponto onde uma estrada atravessa o manguezal.
Segundo Moscatelli et al.(1993), esta estrada, com 1800 m de extensão, foi construída quase que integralmente sobre o manguezal, pela Empresa Inicial Engenharia Ltda em 1987 para ligar a rodovia BR-101 ao Condomínio da Ilha do Jorge. Esta obra em função da ausência de manilhas ou de qualquer outro mecanismo que permitisse a circulação das águas no manguezal afetou profundamente este ecossistema.
Durante o trabalho expedito para elaboração do presente relatório não foram observados indivíduos de Rhizophora mangle de grande porte conforme os descritos por Moscatelli et al. (1993). A ausência destes exemplares confirma o impacto ambiental causado a este manguezal pela construção da estrada. Outra corroboração deste dano é a ocorrência de uma grande quantidade de indivíduos de Hibiscus pernambucensis (algodoeiro da praia) e Acrostichum aureum (avencão), em diversos trechos do manguezal localizados próximos à estrada, que segundo o sistema de classificação para feições de manguezal proposto por Pires (1992) é uma característica de mangues alterados. Sendo assim, pelo acima exposto, pode-se afirmar que a construção desta estrada foi um fator decisivo na modificação da paisagem e da estrutura do manguezal da ilha do Jorge.
As espécies de mangue que ocorreram na parcela 5 foram Rhizophora mangle, Laguncularia racemosa e Avicennia schaueriana. O número de exemplares mensurados foi 43 que apresentaram variação de 2,6 m a 7,5 m de altura. A altura média obtida para esta parcela foi de 5,4 m como apresentado na Tabela 18. Nesta parcela também foi evidenciada a presença de corte de troncos de árvores, porém sobre a espécie Laguncularia racemosa. A espécie de maior contribuição para a área basal foi Rhizophora mangle (Tabela 19). O DAP médio calculado para esta parcela foi de 6,68 cm e a classe diamétrica de maior contribuição para área basal foi a do DAP ³ 5 e < 10 com 0,124242 m2/0,01 ha.
Os dados quantitativos apresentados na Tabela 18 estão representados na Figura 16, onde pode-se observar a dominância de Rhizophora mangle em todas as parcelas estudadas para a ilha do Jorge. Os dados da Tabela 19 indicaram que a classe que mais contribuiu para a área basal total foi do DAP ³ 10, com valor total de 0,61339 m2/0,01 ha, que permitiu caracterizar uma maturidade estrutural do bosque deste perfil.
Figura 16 - Composição quantitativa das espécies nas parcelas amostrais do manguezal da Ilha do Jorge.
7.1.4.4. Manguezal do Ariró - topo
Representa a maior formação deste ecossistema na região de Angra dos Reis, estendendo-se desde o contato com o mar até o trecho superior da Estrada Rio - Santos. Este ambiente é formado principalmente nas bacias de drenagem dos rios Ariró, Floresta e Florestão. O acesso a este manguezal pode ser feito através do mar e dos rios ou através de uma estrada de terra que se comunica com a rodovia Rio - Santos e localiza-se no lado esquerdo (no sentido Angra-Parati) do rio Florestão. Apesar de toda sua exuberância o manguezal do Ariró, a exemplo de outros manguezais brasileiros, está sofrendo os reflexos da ação antrópica. É possível observar trechos degradados por desmatamento, lançamento de lixo e esgoto.
A comunidade que freqüenta o Manguezal do Ariró da Praia do Recife é formada basicamente por pescadores profissionais e ocasionais. Além dos peixes, os mariscos e os caranguejos são intensamente capturados neste manguezal. Os peixes são capturados com auxílio de tarrafas, enquanto que os mariscos são capturados manualmente durante a maré baixa e transportados no momento de maré cheia com auxílio de canoas. A captura dos caranguejos é feita através do tamponamento e do laço e é facilitada pela predominância de um sedimento com características areno-lamosas.
O perfil amostral foi posicionado no trecho do manguezal localizado ao lado da margem direita do rio Florestão, onde foram dispostas 6 parcelas amostrais totalizando 300 m de perfil. As coordenadas geográficas das parcelas estão apresentadas na Tabela 13.
A parcela 1 foi caracterizada através da amostragem de 35 exemplares, onde observou-se a ocorrência de Rhizophora mangle, Laguncularia racemosa e Avicennia schaueriana. Foram observados nesta parcela indivíduos de Rhizophora mangle com os troncos cortados. A altura variou entre 2,8 m e 9,5 m, com média de 5,91 m (Tabela 20).
Tabela 20 - Número de indivíduos e valores de altura obtidos nas amostragens realizadas no perfil do Manguezal do Ariró.
Parcela |
N |
Altura do bosque (m) |
| Superior |
Inferior |
Média |
| 1 |
35 |
9,5 |
2,8 |
5,91 |
| 2 |
31 |
9 |
2,8 |
5,60 |
| 3 |
21 |
10,5 |
3,8 |
7,31 |
| 4 |
24 |
10 |
3,1 |
6,21 |
| 5 |
25 |
8 |
2,7 |
5,39 |
| 6 |
81 |
7,5 |
2,5 |
4,09 |
A espécie de maior contribuição para área basal nesta parcela foi Laguncularia racemosa (Tabela 21). O DAP médio obtido foi de 7,82 cm e a classe diamétrica que mais contribuiu para a área basal foi a do DAP ³ 10 com 0,134186 m2/0,01 ha.
Tabela 21 - Dados de estrutura relacionados a área basal no perfil do Manguezal do Ariró.
P |
N.
ind |
DAP
(médio) |
Rm |
Lr |
As |
Total |
< 2,5 |
>2,5 e < 5 |
> 5 e < 10 |
>10 |
1 |
35 |
7,82 |
0,043956 |
0,109714 |
0,072904 |
0,226574 |
0,000485 |
0,009273 |
0,082629 |
0,134186 |
2 |
31 |
8,36 |
0,074617 |
0,071068 |
0,098234 |
0,243919 |
0,000000 |
0,006708 |
0,088418 |
0,148793 |
3 |
21 |
12,27 |
0,121244 |
0,061404 |
0,220227 |
0,402875 |
0,000000 |
0,002248 |
0,041782 |
0,358845 |
4 |
24 |
8,07 |
0,028376 |
0,051949 |
0,105703 |
0,186028 |
0,000000 |
0,012410 |
0,057926 |
0,115692 |
5 |
25 |
9,52 |
0,108518 |
0,050514 |
0,035857 |
0,194888 |
0,000000 |
0,002138 |
0,088894 |
0,103857 |
6 |
81 |
3,31 |
0,000000 |
0,074280 |
0,048709 |
0,122989 |
0,012110 |
0,069050 |
0,017758 |
0,024072 |
Total |
217 |
8,225 |
0,376710 |
0,418929 |
0,581634 |
1,377273 |
0,012595 |
0,101826 |
0,377406 |
0,885445 |
OBS.: P - Parcela; Rm - Rhizophora mangle; Lr - Laguncularia racemosa; As - Avicennia schaeuriana.
Na parcela 2, a composição específica foi representada por Rhizophora mangle, Laguncularia racemosa e Avicennia schaueriana, totalizando 31 exemplares amostrados. Nesta parcela foram observados vários indivíduos de Laguncularia racemosa tombados. A altura média do bosque foi de 5,60 m e a altura dos espécimes variou entre 2,8 m e máximo de 9 m.
O DAP médio obtido foi de 8,36 cm. A classe diamétrica de maior contribuição para área basal foi a do DAP ³ 10 com 0,148793 m2/0,01 ha. A espécie que mais contribuiu com a área basal nesta parcela foi Avicennia schaueriana (Tabela 21).
A parcela 3 foi representada por 21 exemplares das espécies Rhizophora mangle, Laguncularia racemosa e Avicennia schaueriana. Nesta última espécie foram registrados os indivíduos de maior altura e maior CAP da parcela. Foram observados indivíduos de Rhizophora mangle cortados. Os valores de registrados para altura do bosque variaram entre 3,8 m e 10,5 m. A altura média obtida foi de 7,31 m (Tabela 20).
A espécie de maior contribuição para área basal foi Avicennia schaueriana seguida por Rhizophora mangle. Com relação à classe diamétrica, a maior contribuição para área basal foi a do DAP ³ 10 com 0,358845 m2/0,01 ha, fato que indica um bom desenvolvimento e maturidade do bosque. O DAP médio registrado nesta parcela foi de 12,27 cm (Tabela 21), que representou o maior valor de DAP médio da parcela.
Nos trechos do manguezal entre as parcelas 3 e 4 foram observados exemplares de Avicennia schaueriana bem altos e desenvolvidos. Em função disso, possivelmente os exemplares desta espécie são os mais antigos deste trecho do manguezal estudado.
Na parcela 4 foi registrada a ocorrência de Rhizophora mangle, Laguncularia racemosa e Avicennia schaueriana. Observou-se a ocorrência de indivíduos de Laguncularia racemosa tombados e cortados. O número de indivíduos amostrados foi de 21 exemplares (Tabela 20). A altura média do bosque foi de 6,21 m, sendo que os valores de altura oscilaram entre 3,1 m e 10 m. O DAP médio obtido para a parcela 4 foi de 8,07 cm (Tabela 21). A classe diamétrica de maior contribuição para área basal foi a do DAP ³ 10 com 0,115692 m2/0,01 ha. A espécie que mais contribuiu foi Avicennia schaueriana para a área basal nesta parcela.
Para a parcela 5 a composição específica do bosque foi representada por Rhizophora mangle, Laguncularia racemosa e Avicennia schaueriana. Foram observados exemplares de Laguncularia racemosa com o tronco cortado. A altura mínima foi de 2,7 m e a máxima de 8 m. O valor médio de altura foi de 5,39 m (Tabela 20) e o número de exemplares amostrados foi de 25. A espécie que mais contribuiu para a área basal desta parcela foi Rhizophora mangle. O DAP médio registrado foi de 9,52 cm (Tabela 21). A classe diamétrica ³ 10 foi a de maior contribuição para área basal com 0,103857 m2/0,01 ha.
A parcela 6 foi a que apresentou maior número de exemplares em todo o perfil com um total de 81 espécimes. A altura do bosque variou entre 2,5 m e 7,5 m. O valor médio registrado para altura foi de 4,09 m que foi o menor de todo o perfil (Tabela 20). As espécies observadas foram Laguncularia racemosa e Avicennia schaueriana. A espécie que apresentou maior contribuição para área basal da parcela 6 foi Laguncularia racemosa. A classe diamétrica de maior contribuição para área basal foi a do DAP ³2,5 e < 5 com 0,069050 m2/0,01 ha. O DAP médio de 3,31 cm foi o menor de todo o perfil (Tabela 21).
Após esta parcela, o bosque do manguezal ainda se distribui até uma distância de aproximadamente mais 30 m, quando este ambiente faz contato com uma mata completamente antropizada. O reflexo da atividade antrópica é visível na porção do manguezal mais próxima a mata onde ocorre um intenso corte de troncos de exemplares jovens de Laguncularia racemosa. A Figura 17 apresenta a contribuição quantitativa de cada espécie ao longo do perfil, onde destaca-se a dominância de Laguncularia racemosa em quase todas as parcelas.
Figura 17 - Composição quantitativa das espécies nas parcelas amostrais do manguezal do Ariró.
Os valores de área basal, demonstrados na Tabela 21, indicaram que a maior contribuição para área basal total foi do DAP ³ 10 com 0,885445 m2/0,06 ha, fato que permitiu caracterizar este perfil como mais maduro estruturalmente. A espécie que mais contribui para a área basal total foi Avicennia schaueriana.
A fauna observada no Manguezal do Ariró por moluscos e crustáceos. Entre os moluscos foi observada a ocorrência de Littorina (Scabra) angulifera (buso do mangue), Teredo spp. (broca da madeira) e Ostrea rhizophorae (ostra do mangue).
Os crustáceos foram representados pelas seguintes espécies: Chthamalus spp. (craca), Balanus spp. (craca), Callinectes spp. (siri), Goniopsis cruentata (aratu vermelho e preto), Aratus pisonii (marinheiro), Eurytium sp. (goiá) Uca (Minuca) rapax, (chama-maré), Uca (Minuca) mordax (chama-maré) Uca (Celuca) uruguayensis (chama-maré), Uca (Celuca) leptodactyla (chama-maré) e Ucides cordatus (caranguejo uçá). Nas parcelas amostrais foi observada uma grande quantidade de armadilhas (laços) para captura de Ucides cordatus, bem como foi registrada a atividade de cata de caranguejos pelos pescadores. Este manguezal também é visitado por catadores de caranguejos de outros municípios que acampam no manguezal para efetuar a pesca destes crustáceos.
7.1.4.5. Manguezal da Pousada do Bosque - topo
Localiza-se na área de influência da drenagem do rio Perequê. Apesar de não ser um manguezal muito extenso este ambiente é bastante representativo na região, pois proporciona subsistência e lazer para a população local. Este manguezal está instalado sobre um sedimento com características arenosas que é proveniente do transporte de partículas feitas pelo rio Perequê. O processo de sedimentação está se acelerando em função da derrubada da mata ciliar e da ocupação das margens do rio Perequê, que se reflete no manguezal.
Apesar de aparentemente ser uma área isolada em virtude da presença do Hotel Pousada do Bosque que está localizado ao seu lado, este manguezal é alvo de uma intensa captura de Ucides cordatus (caranguejo uçá) e Cardisoma guanhumi (guaiamum) por pessoas da comunidade do entorno que penetram neste manguezal pela estrada Rio - Santos.
O perfil amostral do manguezal da Pousada do Bosque foi posicionado na margem direita do rio Perequê, próximo a sua foz. Foram efetuadas amostragens em três parcelas deste manguezal. As coordenadas geográficas de cada parcela estão apresentadas na Tabela 13.
Na parcela 1 a composição específica do bosque foi representada apenas por Laguncularia racemosa. Próximo a esta parcela foram observados indivíduos desta espécie mortos por deposição de areia. O número total de indivíduos amostrados foi de 37 (Tabela 22). A altura média do bosque foi de 5,83 m, sendo que os exemplares apresentaram a altura mínima de 2,8 m e a máxima de 9,3 m.
Tabela 22 - Número de indivíduos e valores de altura obtidos nas amostragens realizadas no perfil do Manguezal da Pousada do Bosque.
Parcela |
N |
Altura do bosque (m) |
| Superior |
Inferior |
Média |
1 |
37 |
9,3 |
2,8 |
5,83 |
2 |
29 |
6,5 |
2,7 |
4,65 |
3 |
49 |
8 |
2,5 |
4,33 |
A classe diamétrica que mais contribuiu para a área basal foi do DAP ³ 5 e < 10 com 0,120132 m2/0,01 ha. O DAP médio registrado foi de 5,98 cm (Tabela 23).
Tabela 23 - Dados de estrutura relacionada a área basal no perfil da Pousada do Bosque.
P |
N.
ind |
DAP
(médio) |
Rm |
Lr |
As |
Total |
< 2,5 |
> 2,5 e < 5 |
> 5 e < 10 |
> 10 |
| 1 |
37 |
5,98 |
0 |
0,242351 |
0 |
0,242351 |
0,003878 |
0,021211 |
0,120132 |
0,097129 |
| 2 |
29 |
6,79 |
0 |
0,302237 |
0 |
0,302237156 |
0,003190 |
0,015605 |
0,069246 |
0,214197 |
| 3 |
49 |
5,26 |
0 |
0,282968 |
0 |
0,282968 |
0,006422 |
0,026047 |
0,074085 |
0,176414 |
| Total |
115 |
6,01 |
0 |
0,827556 |
0 |
0,827556 |
0,013490 |
0,062863 |
0,263462 |
0,487740 |
OBS.: P - Parcela; Rm - Rhizophora mangle; Lr - Laguncularia racemosa; As - Avicennia schaeuriana.
Na parcela 2, a altura do bosque variou entre 2,7 m e 6,5 m. O valor médio registrado para a altura dos exemplares foi de 4,65 m (Tabela 22). Nesta parcela também foi observada somente a ocorrência de Laguncularia racemosa. Próximo esta parcela também foi registrada a mortandade por soterramento de exemplares desta espécie. O DAP médio obtido nesta parcela foi de 6,79 cm conforme apresentado na Tabela 23. A classe diamétrica de DAP ³ 5 e < 10 com 0,069246 m2/0,01 ha foi a que mais contribuiu para a área basal.
O bosque da parcela 3 também foi monoespecífico com ocorrência de Laguncularia racemosa. O número de exemplares amostrados foi de 49, o maior deste perfil. A altura do bosque variou entre 2,5 m e 8 m. O valor médio registrado para altura foi de 4,33 m, que foi o menor valor registrado para o perfil (Tabela 22). O DAP médio obtido na parcela 3 foi de 5,23 cm (Tabela 23). A classe diamétrica de maior contribuição foi a do DAP ³ 10 com 0,176414 m2/0,01 ha.
Os dados quantitativos apresentados na Tabela 23 estão representados na Figura 18, onde pode-se observar a dominância de Laguncularia racemosa em todas as parcelas estudadas.
Figura 18 - Composição quantitativa das espécies nas parcelas amostrais do manguezal da Pousada do Bosque.
Os dados da Tabela 23 indicaram que a classe que mais contribuiu para a área basal total foi do DAP ³ 10, com valor total de 0,487740 m2/0,03 ha.
A fauna deste perfil foi representada por moluscos e crustáceos. Entre a malacofauna foram observadas as seguintes espécies: Littorina (Scabra) angulifera (buso do mangue), Anomalocardia brasiliana (berbigão), Ostrea rhizophorae (ostra do mangue), Lucina pectinata (ameixa), Macoma constricta (tarioba) e Mytella spp.(sururu). A captura de moluscos é feita tanto na franja do manguezal quanto no interior do bosque. No interior do bosque estes animais são capturados principalmente nos canais que drenam este ambiente. Em diversos pontos foram observados depósitos de conchas de moluscos, principalmente de Lucina pectinata, que após a captura são fervidos e descarnados dentro do manguezal. As conchas são depositadas em trechos de sedimento mais duro onde predomina Hibiscus pernambucensis. Nestas áreas também é possível observar restos de fogueiras e as latas utilizadas para o transporte dos animais. A presença de grandes bancos de Mytella spp. próximos aos canais de drenagem constitui um importante item na subsistência das comunidades pesqueiras locais. Cabe ainda ressaltar que os bancos destes animais são raros nos manguezais do Estado do Rio de Janeiro e estão desaparecendo em função da poluição.
Os representantes da carcinofauna foram: Chthamalus spp. (craca), Balanus spp. (craca), Uca spp. (chama-maré), Aratus pisonii (marinheiro), Goniopsis cruentata (aratu vermelho e preto), Ucides cordatus (caranguejo uçá) e Cardisoma guanhumi (guaiamum). Ao longo de todo o perfil foi possível observar a presença armadilhas para captura de caranguejo, indicando que este manguezal também é muito explorado por caranguejeiros. Nas tocas de Ucides cordatus foram observados os laços e nas áreas de sedimento mais duro, onde ocorrem Hibiscus pernambucensis e as tocas de Cardisoma guanhumi, observou-se a presença das armadilhas conhecidas como “ratoeiras”, próprias para a captura de guaiamum.
7.1.4.6. Avaliação Geral das Áreas Amostradas - topo
Os resultados obtidos demonstraram que, de um modo geral, os bosques estudados nos três perfis apresentaram predominância de Rhizophora mangle (mangue sapateiro) onde observou-se 454 exemplares, enquanto que ocorreram 453 exemplares de Laguncularia racemosa (mangue branco) e 85 exemplares de Avicennia schaueriana (mangue preto). Os totais observados de cada espécie estão apresentados na Tabela 24.
Tabela 24 - Numero total de indivíduos encontrados por espécie nas áreas amostradas.
Perfis |
Rhizophora mangle |
Laguncularia racemosa |
Avicennia schaueriana |
Número de indivíduos |
| Bracuí |
66 |
123 |
0 |
189 |
| Praia do Recife |
223 |
69 |
16 |
308 |
| Ilha do Jorge |
121 |
25 |
17 |
163 |
| Ariró |
44 |
121 |
52 |
217 |
| Pousada do Bosque |
0 |
115 |
0 |
115 |
| Total |
454 |
453 |
85 |
992 |
A partir dos dados da tabela acima, pôde-se montar um gráfico da distribuição das espécies que demonstra a composição de cada bosque avaliado (Figura 19).
Figura 19 - Numero de indivíduos encontrados por espécie nas áreas amostradas.
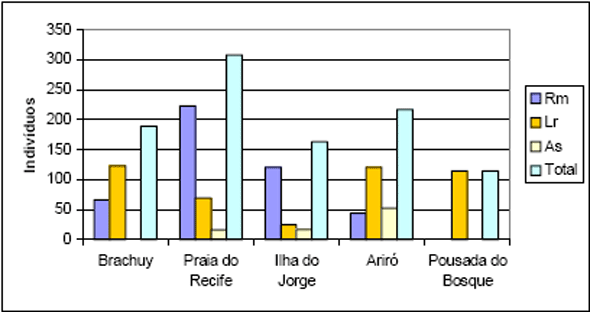
Rm - Rhizophora mangle; Lr - Laguncularia racemosa; As - Avicennia schaueriana.
A espécie Rhizophora mangle apresentou maior representatividade em termos quantitativos nos perfis da Praia do Recife e da Ilha do Jorge, enquanto os perfis do Bracuí, Ariró e Pousada do Bosque apresentaram maior contribuição de Laguncularia racemosa. Estes dados indicaram que os manguezais dos perfis do Bracuí, Ariró e Pousada do Bosque foram representados por um bosque do tipo branco, enquanto que os manguezais dos perfis da Praia do Recife e da Ilha do Jorge foram representados por um bosque do tipo vermelho.
Com base nos dados obtidos para o total das parcelas amostradas pode-se observar que Rhizophora mangle (Rm) apresentou um percentual de ocorrência de 62%, seguido por Laguncularia racemosa com 34% e Avicennia schaueriana com 4% (Figura 20).
Figura 20 - Percentual de ocorrência das espécies no total das áreas estudadas.
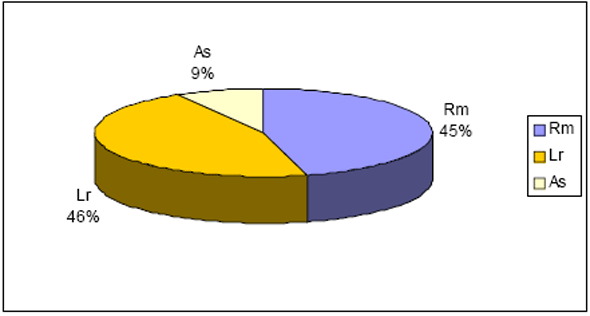
A partir dos dados obtidos no campo pôde-se observar que a maior densidade de 44.000 ind/ha ocorreu no perfil do manguezal da Praia do Recife, enquanto a menor densidade foi de 23.625 ind/ha registrada no perfil do manguezal do Bracuí (Tabela 25).
Tabela 25 - Numero de indivíduos e valores médios de DAP e altura nos perfis estudados.
| Perfis |
Ind. Amostrados |
Área
Amostrada |
N. Ind./ m2 |
N.Ind.
(ha) |
DAP
(média) |
Altura
(média) |
| Bracuí |
189 |
8 x 10m |
2,36 |
23.625 |
5,90 |
5,01 |
| Praia do Recife |
308 |
7 x 10m |
4,40 |
44.000 |
6,35 |
5,17 |
| Ilha do Jorge |
163 |
5 x 10m |
3,26 |
32.600 |
8,06 |
6,33 |
| Ariró |
217 |
6 x 10m |
3,62 |
36.167 |
8,23 |
5,75 |
| Pousada do Bosque |
115 |
3 x 10m |
3,83 |
38.333 |
6,01 |
4,94 |
Os maiores valores médios de DAP e altura foram observados no perfil do manguezal da ilha do Jorge. O menor valor de DAP médio foi registrado no perfil do manguezal do Bracuí e o menor valor para altura média foi mensurado no perfil do manguezal da Pousada do Bosque.
Os valores de contribuição de cada espécie e de cada classe diamétrica para a área basal estão apresentados na
Tabela 26 e corroboram os resultados já demonstrados anteriormente para a dominância de espécies, onde Rhizophora mangle apresentou a maior contribuição para área basal de 2,43 m2/0,15 ha, sendo também a espécie com maior número de exemplares e conseqüentemente o maior percentual de freqüência nas parcelas estudadas.
A classe diamétrica que mais contribuiu para a área basal total foi a do DAP ³ 10 cm com total de 2,99 m2/0,15 ha, seguida pela classe ³ 5 e < 10 cm que totalizou 1,77 m2/0,15 ha. Estes dados indicam que de um modo geral os bosques estudados apresentaram um bom desenvolvimento estrutural. Entretanto cabe ressaltar que alguns trechos do perfil do manguezal do Bracuí e da ilha do Jorge encontram-se em franco processo de recuperação.
A partir dos dados coletados em campo foram calculados os valores médios para DAP e altura de cada parcela. Estes resultados estão apresentados na Tabela 27. Pode-se concluir que de um modo geral os bosques estudados não são altos. A exceção fica por conta da parcela 2 do perfil do manguezal da ilha do Jorge onde registrou-se uma altura média de 9,46 m. O menor valor de altura média foi obtido na parcela 1 do manguezal da praia do Recife com 3,38 m. Com relação ainda a altura houve semelhanças estruturais entre: as parcelas 4, 6 e 7 no manguezal do Bracuí, as parcela 1 e 2 e 3 e 4 no manguezal da praia do Recife e entre as parcelas 3 e 4 no manguezal da ilha do Jorge.
O maior valor médio para DAP foi registrado na parcela 3 do perfil do manguezal do Ariró, enquanto que o menor valor médio foi obtido na parcela 1 do manguezal do Bracuí. Comparando os cinco perfis estudados, os maiores valores médios de DAP foram observados no manguezal da ilha do Jorge. Os valores médios de DAP apresentaram semelhança entre: as parcelas 7 e 8 do manguezal do Bracuí, as parcelas 1 e 2 e 3 e 4 do manguezal da praia do Recife e as parcelas 2 e 3 do manguezal da ilha do Jorge.
Quando se compararam os dados de DAP e de altura com os resultados de densidade, pôde-se observar que existiu uma coincidência entre os maiores valores médios com o trecho de estágio estrutural mais avançado (perfil da ilha do Jorge), pois ocorre uma redução do número de exemplares por hectare.
Tabela 26 - Contribuição de cada espécie e de cada classe diamétrica para a área basal (m2/0,01 ha) nos perfis estudados.
Perfis |
Área basal das espécies |
Área basal pelas classes de DAP |
| Rm |
Lg |
As |
Total |
< 2,5 cm |
> 2,5 e
< 5 cm |
> 5 e < 10 cm |
> 10 cm |
| Bracuí |
0,397171 |
0,277705 |
- |
0,674875 |
0,00387 |
0,088144 |
0,366816 |
0,216045 |
| Praia do Recife |
1,014026 |
0,234699 |
0,16828 |
1,417005 |
0,013267 |
0,161891 |
0,452909 |
0,788936 |
| Ilha do Jorge |
0,646995 |
0,129617 |
0,187389 |
0,964001 |
0,009445 |
0,035423 |
0,305742 |
0,613391 |
| Ariró |
0,37671 |
0,418929 |
0,581634 |
1,377273 |
0,012595 |
0,101826 |
0,377406 |
0,885445 |
| Pousada do Bosque |
- |
0,827556 |
- |
0,827556 |
0,01349 |
0,062863 |
0,263462 |
0,48774 |
| Total |
2,434902 |
1,888506 |
0,937303 |
5,26071 |
0,052667 |
0,450147 |
1,766335 |
2,991557 |
Rm - Rhizophora mangle; Lr - Laguncularia racemosa; As - Avicennia schaeuriana.
Tabela 27 - Valores médios de DAP (cm) e Altura (m) dos espécimes do manguezal do Bracuí, Praia do Recife, Ilha do Jorge, Ariró e Pousada do Bosque.
Parcela |
Bracuí |
Praia do Recife |
Ilha do Jorge |
Ariró |
Pousada do Bosque |
DAP |
Altura |
DAP |
Altura |
DAP |
Altura |
DAP |
Altura |
DAP |
Altura |
| Parcela 1 |
3,18 |
3,88 |
7,13 |
5,65 |
3,70 |
3,97 |
7,82 |
5,91 |
5,98 |
5,83 |
| Parcela 2 |
* |
* |
7,12 |
6,35 |
11,14 |
9,46 |
8,36 |
5,60 |
6,79 |
4,65 |
| Parcela 3 |
* |
* |
4,04 |
3,38 |
10,31 |
6,29 |
12,27 |
7,31 |
5,26 |
4,33 |
| Parcela 4 |
4,17 |
5,44 |
4,13 |
4,96 |
8,46 |
6,53 |
8,07 |
6,21 |
- |
- |
| Parcela 5 |
8,75 |
6,69 |
7,73 |
5,85 |
6,68 |
5,40 |
9,52 |
5,39 |
- |
- |
| Parcela 6 |
5,75 |
5,13 |
6,44 |
4,64 |
- |
- |
3,31 |
4,09 |
- |
- |
| Parcela 7 |
6,70 |
4,93 |
7,86 |
5,33 |
- |
- |
- |
- |
- |
- |
| Parcela 8 |
6,82 |
4,01 |
- |
- |
- |
- |
- |
- |
- |
- |
* - sem indivíduos dentro do critéruio de amostragem
Os dados apresentados na Tabela 27 foram também organizados graficamente, de modo a permitir observar a variação estrutural nas parcelas ao longo de cada perfil estudado, e estão apresentados a seguir.
Na Figura 21, pode-se observar que de maneira geral os valores médios de DAP crescem no sentido da parcela 1 para a parcela 8. O valor médio máximo registrado para esta variável ocorreu na parcela 5 e o valor médio mínimo na parcela 1.
Os valores médios de altura foram crescentes até a parcela 5, decrescendo a partir daí. O valor médio máximo foi observado na parcela 5 e o mínimo na parcela 1. A parcela mais desenvolvida estruturalmente foi a parcela 5.
Os dados indicaram uma forte correlação entre DAP e altura, tanto para os valores médios máximos presentes na parcela 5, quanto para os valores médios mínimos registrados na parcela 1.
Figura 21 - Variação dos valores médios de DAP (cm) e altura (m) ao longo do perfil do manguezal do Bracuí.
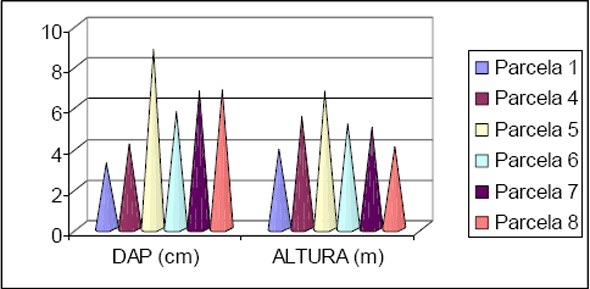
A variação dos valores médios observados no perfil do manguezal da praia do Recife está demonstrada na Figura 22.
Neste perfil, a variação dos valores de DAP foi bem diferenciada. Os maiores valores médios foram observados nas parcelas 5 e 7, enquanto que os menores valores médios foram obtidos nas parcelas 3 e 4. O mesmo ocorreu com relação a altura. As parcelas 1 e 2 foram as que apresentaram melhor desenvolvimento estrutural e a parcela 3 apresentou o menor desenvolvimento estrutural.
Na Figura 23 está demonstrada a variação dos valores médios de DAP e altura obtidos no perfil do manguezal da Ilha do Jorge.
Figura 22 - Variação dos valores médios de DAP (cm) e altura (m) ao longo do perfil do manguezal da Praia do Recife.
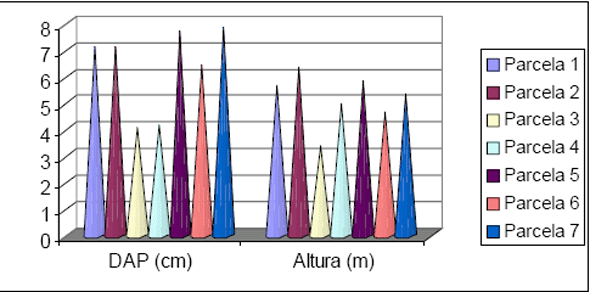
Figura 23 - Variação dos valores médios de DAP (cm) e altura (m) ao longo do perfil do manguezal da Ilha do Jorge.
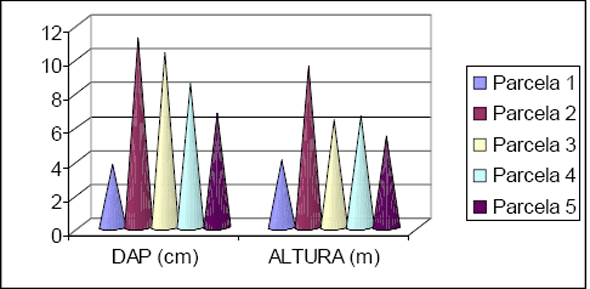
Os valores médios de DAP aumentaram da parcela 1 para a parcela 2, decrescendo a partir daí. O valor médio máximo foi observado na parcela 2 e o valor médio mínimo na parcela 1.
Observando-se os dados constatou-se que existe uma semelhança entre a parcela 2 e parcela 3.
Os valores de altura apresentaram-se bastante variados. O valor médio mínimo foi observado na parcela 1, enquanto que o valor médio máximo foi obtido na parcela 2. Os dados indicaram uma semelhança entre as parcelas 3 e 4. A parcela 2 foi a que apresentou melhor desenvolvimento estrutural. Conforme observado no perfil do manguezal do Bracuí, no perfil do manguezal da Ilha do Jorge os valores médios máximos e mínimos de DAP e altura também apresentaram uma forte correlação, ocorrendo respectivamente nas parcelas 2 e 1.
Figura 24 - Variação dos valores médios de DAP (cm) e altura (m) ao longo do perfil do manguezal da praia do Recife.
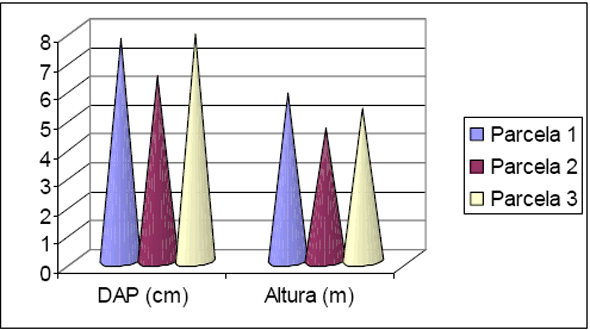
Na Figura 24, pode-se observar pelos valores de DAP médio e altura que estruturalmente as parcelas 1 e 3 são bem semelhantes. Os valores mínimos para estas variáveis foram registrados na parcela 2 que conseqüentemente possui uma condição estrutural inferior as parcelas 1 e 3. A variação dos valores médios de altura e DAP obtidos no perfil do manguezal do Ariró está demonstrada na Figura 25.
Figura 25 - Variação dos valores médios de DAP (cm) e altura (m) ao longo do perfil do manguezal do Ariró.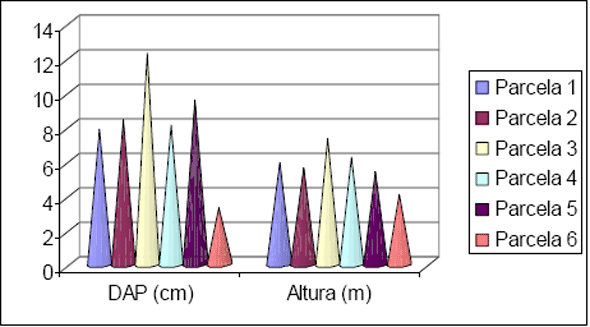
Neste perfil, a variação dos valores de DAP médio foi bem diferenciada. O maior valor foi observado na parcela 3 e o menor na parcela 6. Comparando-se os dados observa-se que existe uma semelhança entre as parcelas 1, 2 e 4. A variação da altura média foi diferente da variação do DAP médio. Ao observar os valores de altura constata-se que existe uma semelhança entre as parcelas 1, 2 e 5. Os maiores valores médios observados nas parcelas 3 e 4 pode ser explicado pela presença de exemplares de grande porte de Avicennia schaueriana e Rhizophora mangle.
Na Figura 26 está demonstrada a variação dos valores médios de DAP e altura obtidos no perfil do manguezal da Pousada do Bosque.
Figura 26 - Variação dos valores médios de DAP (cm) e altura (m) ao longo do perfil do manguezal da Pousada do Bosque.
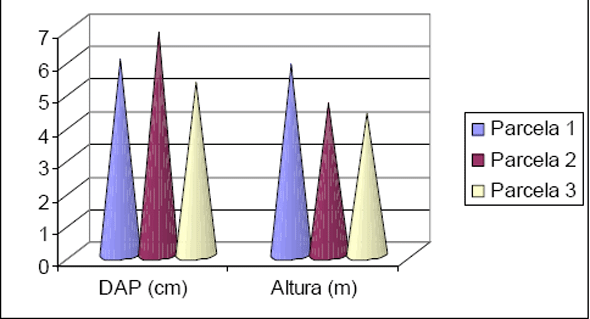
Os valores médios de DAP aumentaram da parcela 1 para a parcela 2, decrescendo na parcela 3. O valor médio máximo foi observado na parcela 2 e o valor médio mínimo na parcela 3.
Os valores de altura apresentaram-se decrescentes da parcela 1 para a parcela 3. O valor médio máximo foi observado na parcela 1, enquanto que o valor médio mínimo foi obtido na parcela 3.
Os dados coletados durante o trabalho expedito foram também utilizados para realizar a análise fitossociológica. Esta análise foi efetuada através de uma abordagem individualizada em perfis, de modo a apresentar as características de cada bosque amostrado.
Os dados da análise fitossociológica do perfil do manguezal do Bracuí estão demonstrados na Tabela 28.
Tabela 28 - Avaliação fitossociológica do bosque no perfil do manguezal do Bracuí.
Bracuí
Parcela |
Ind |
Troncos |
Área basal |
| Rm |
Lr |
As |
Rm |
Lr |
As |
Rm |
Lr |
As |
| 1 |
0 |
37 |
0 |
0 |
58 |
0 |
|
0,052042 |
0 |
| 4 |
1 |
31 |
0 |
1 |
52 |
0 |
0,003993 |
0,075577 |
0 |
| 5 |
20 |
6 |
0 |
20 |
6 |
0 |
0,130499 |
0,043699 |
0 |
| 6 |
13 |
28 |
0 |
13 |
46 |
0 |
0,090745 |
0,094941 |
0 |
| 7 |
12 |
9 |
0 |
12 |
11 |
0 |
0,068573 |
0,025641 |
0 |
| 8 |
20 |
12 |
0 |
20 |
13 |
0 |
0,103361 |
0,037845 |
0 |
| Soma |
66 |
123 |
0 |
66 |
186 |
0 |
0,397171 |
0,277705 |
0 |
| Total |
|
189 |
|
|
252 |
|
|
0,674876 |
|
| Espécie |
Parcelas |
Ind. |
AB |
DR |
FA |
FR |
DoR |
IVI |
IVC |
| R. mangle |
5 |
66 |
0,39 |
0,35 |
83,3 |
45,44 |
0,59 |
46,38 |
0,94 |
| L.racemosa |
6 |
123 |
0,27 |
0,65 |
100 |
54,56 |
0,41 |
55,62 |
1,06 |
| A schaueriana |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
| |
6 |
189 |
0,66 |
1 |
183,3 |
100 |
1 |
|
|
Rm - Rhizophora mangle; Lr - Laguncularia racemosa; As - Avicennia schaueriana; AB - Área basal; DR - Densidade relativa; FA - Freqüência absoluta; FR - Freqüência relativa; DoR - Dominância relativa; IVI - Índice de valor de importância; IVC - Índice de valor de cobertura.
A análise dos dados revelou a presença de uma grande quantidade de exemplares nas Parcelas. Nas Parcelas 1, 4 e 6 pode-se observar um valor muito elevado de troncos de Laguncularia racemosa em relação aos indivíduos, demonstrando um estresse gerado no bosque pelo corte de madeira que propiciou o surgimento de vários exemplares ramificados neste taxa. Apesar disso, Rhizophora mangle foi a espécie que apresentou neste perfil maior contribuição para área basal total. Cabe ainda destacar que nas parcelas estudadas não foi observado nenhum exemplar de Avicennia schaueriana.
Os resultados apresentados na Tabela 28 indicaram que para o número de indivíduos (Ind.), densidade relativa (DR), freqüência absoluta (FA), freqüência relativa (FR) índice de valor de importância (IVI) e índice de valor de cobertura (IVC) os valores de Laguncularia racemosa foram superiores aos valores de Rhizophora mangle. Porém, os valores observados para área basal (AB) e dominância relativa (DoR) foram superiores para Rhizophora mangle. A fisiografia deste perfil é de manguezal de franja. Os valores obtidos na análise fitossociológica corroboram a avaliação da estrutura do bosque realizada neste perfil.
Tabela 29 - Avaliação fitossociológica do bosque no perfil do manguezal da Praia do Recife.
Praia do Recife Parcelas |
Ind |
Troncos |
Área basal |
| Rm |
Lr |
As |
Rm |
Lr |
As |
Rm |
Lr |
As |
| 1 |
17 |
10 |
4 |
22 |
15 |
7 |
0,219457 |
0,036430 |
0,003561 |
| 2 |
19 |
15 |
1 |
20 |
23 |
3 |
0,212219 |
0,036236 |
0,023469 |
| 3 |
31 |
5 |
0 |
31 |
6 |
0 |
0,040306 |
0,016433 |
0 |
| 4 |
84 |
6 |
1 |
85 |
6 |
1 |
0,109642 |
0,019656 |
0,019894 |
| 5 |
17 |
16 |
7 |
17 |
19 |
9 |
0,185223 |
0,024126 |
0,118615 |
| 6 |
36 |
14 |
3 |
37 |
15 |
4 |
0,139808 |
0,094682 |
0,00274 |
| 7 |
19 |
3 |
0 |
19 |
3 |
0 |
0,107371 |
0,007138 |
0 |
| Soma |
223 |
69 |
16 |
231 |
87 |
24 |
1,014026 |
0,234701 |
0,168279 |
| Total |
|
308 |
|
|
342 |
|
|
1,417006 |
|
| Espécie |
Parcelas |
Ind |
AB |
DR |
FA |
FR |
DoR |
IVI |
IVC |
| R. mangle |
7 |
223 |
1,014 |
0,72 |
100 |
0,37 |
0,716 |
1,81 |
1,73 |
| L.racemosa |
7 |
69 |
0,235 |
0,22 |
100 |
0,37 |
0,166 |
0,76 |
0,40 |
| A schaueriana |
5 |
16 |
0,168 |
0,05 |
71 |
0,26 |
0,119 |
0,43 |
0,29 |
| |
7 |
308 |
1,417 |
1,00 |
271 |
100 |
1 |
|
|
Rm- Rhizophora mangle; Lr - Laguncularia racemosa; As - Avicennia schaueriana; AB - Área basal; DR - Densidade relativa; FA - Freqüência absoluta; FR - Freqüência relativa; DoR - Dominância relativa; IVI - Índice de valor de importância; IVC - Índice de valor de cobertura.
Neste perfil foram observados elevados números de indivíduos, principalmente na Parcela 4. O maior número de troncos em relação ao número de indivíduos observados nas demais parcelas indicaram que as árvores das três espécies, Rhizophora mangle, Laguncularia racemosa e Avicennia schaueriana, apresentaram-se bem ramificadas, demonstrando a ocorrência de estresse gerado por corte de madeira, que foi corroborado pelas observações realizadas no campo.
A maior contribuição para a área basal em todas Parcelas foi da espécie Rhizophora mangle, que conseqüentemente apresentou maior contribuição para a área basal total.
De acordo com os dados da Tabela 29 Rhizophora mangle também apresentou valores superiores de número de indivíduos (Ind.), área basal (AB), densidade relativa (DR), dominância relativa (DoR), índice de valor de importância (IVI) e índice de valor de cobertura (IVC) do que as outras duas espécies. A espécie Laguncularia racemosa apresentou uma importante contribuição para freqüência absoluta (FA) e freqüência relativa (FR) com valores iguais aos registrados para Rhizophora mangle.
Neste bosque a fisiografia é de manguezal de franja. Os resultados obtidos para análise fitossociológica corroboraram a avaliação da estrutura deste perfil.
A análise fitossociológica do bosque do perfil do manguezal da Ilha do Jorge está apresentada na Tabela 30.
Tabela 30 - Avaliação fitossociológica do bosque no perfil do manguezal da Ilha do Jorge.
Ilha do Jorge
Parcela |
Ind |
Troncos |
Área basal |
| Rm |
Lr |
As |
Rm
|
Lr |
As |
Rm |
Lr |
As |
| 1 |
39 |
4 |
6 |
45
|
4 |
10 |
0,043286 |
0,001904 |
0,071874 |
| 2 |
19 |
0 |
2 |
20
|
0 |
2 |
0,210617 |
- |
0,029220 |
| 3 |
15 |
3 |
2 |
15
|
3 |
2 |
0,135582 |
0,020547 |
0,061601 |
| 4 |
19 |
8 |
3 |
19
|
8 |
5 |
0,129592 |
0,064016 |
0,019894 |
| 5 |
29 |
10 |
4 |
29
|
11 |
5 |
0,127919 |
0,043150 |
0,004800 |
| Soma |
121 |
25 |
17 |
128
|
26 |
24 |
0,646995 |
0,129617 |
0,187389 |
| Total |
|
163 |
|
|
178 |
|
|
0,964001 |
|
| Espécie |
Parcelas |
Ind. |
AB |
DR
|
FA |
FR |
DoR |
IVI |
IVC |
| R. mangle |
5 |
121 |
0,64 |
0,74
|
100 |
35,71 |
0,67 |
37,12 |
1,41 |
| L.racemosa |
4 |
25 |
0,13 |
0,16
|
80 |
28,58 |
0,14 |
28,88 |
0,3 |
| A schaueriana |
5 |
17 |
0,18 |
0,1
|
100 |
35,71 |
0,19 |
36,00 |
0,29 |
| |
5 |
163 |
0,95 |
1
|
280 |
100 |
1 |
|
|
Rm - Rhizophora mangle; Lr - Laguncularia racemosa; As - Avicennia schaueriana; AB - Área basal; DR - Densidade relativa; FA - Freqüência absoluta; FR - Freqüência relativa; DoR - Dominância relativa; IVI - Índice de valor de importância; IVC - Índice de valor de cobertura.
Os dados apresentados na Tabela 30 indicam que o número de troncos foi superior ao de indivíduos. Este fato foi registrado principalmente na Parcela 1, indicando a ocorrência de estresse do bosque causado pelo corte de madeira de exemplares de Rhizophora mangle e Avicennia schaueriana, dano que foi confirmado pelas observações realizadas no campo.
A espécie Rhizophora mangle apresentou maior contribuição para a área basal na maioria das Parcelas estudadas. A exceção ocorreu na Parcela 1 onde a maior contribuição foi feita pela espécie Avicennia schaueriana. Sendo assim, Rhizophora mangle foi a espécie que mais contribuiu para a área basal total do perfil.
Os resultados indicaram que os valores registrados para Rhizophora mangle foram superiores quando comparados aos valores das outras espécies para número de indivíduos (Ind.), área basal (AB), densidade relativa (DR), freqüência relativa (FR), dominância relativa (DoR), índice de valor de importância (IVI) e índice de valor de cobertura (IVC). A espécie Avicennia schaueriana apresentou o mesmo valor da espécie Rhizophora mangle para freqüência absoluta (FA).
A fisiografia do bosque do perfil da Ilha do Jorge também é de manguezal de franja. Os dados obtidos para a análise fitossociológica corroboram as observações da avaliação da estrutura do bosque.
A Tabela 31 demonstra os dados da análise fitossociológica do perfil do manguezal do Ariró.
Tabela 31 - Avaliação fitossociológica do bosque no perfil do manguezal do Ariró.
Ariró
Parcela |
Ind |
Troncos |
Área basal |
| Rm |
Lr |
As |
Rm |
Lr |
As |
Rm |
Lr |
As |
| 1 |
10 |
17 |
8 |
10 |
18 |
9 |
0,043956 |
0,109714 |
0,072904 |
| 2 |
9 |
17 |
5 |
9 |
17 |
5 |
0,074617 |
0,071068 |
0,098234 |
| 3 |
9 |
9 |
3 |
10 |
11 |
4 |
0,121244 |
0,061404 |
0,220227 |
| 4 |
2 |
15 |
7 |
2 |
17 |
9 |
0,028376 |
0,051949 |
0,105703 |
| 5 |
14 |
7 |
4 |
14 |
7 |
4 |
0,108518 |
0,050514 |
0,035857 |
| 6 |
0 |
56 |
25 |
0 |
75 |
34 |
0 |
0,074280 |
0,048709 |
| Soma |
44 |
121 |
52 |
45 |
145 |
65 |
0,376710 |
0,418929 |
0,581634 |
| Total |
|
217 |
|
|
255 |
|
|
1,377273 |
|
| Espécie |
Parcelas |
Ind |
AB |
DR |
FA |
FR |
DoR |
IVI |
IVC |
| R. mangle |
5 |
44 |
0,38 |
0,21 |
83,3 |
29,4 |
0,27 |
29,88 |
0,48 |
| L.racemosa |
6 |
121 |
0,42 |
0,56 |
100 |
35,3 |
0,31 |
36,17 |
0,87 |
| A schaueriana |
6 |
52 |
0,58 |
0,23 |
100 |
35,3 |
0,42 |
35,95 |
0,65 |
| |
6 |
217 |
1,38 |
1 |
283,3 |
100 |
1 |
|
|
Rm - Rhizophora mangle; Lr - Laguncularia racemosa; As - Avicennia schaueriana; AB - Área basal; DR - Densidade relativa; FA - Freqüência absoluta; FR - Freqüência relativa; DoR - Dominância relativa; IVI - Índice de valor de importância; IVC - Índice de valor de cobertura.
Neste perfil a espécie que apresentou maior total de indivíduos foi Laguncularia racemosa. Analisando os dados de cada parcela esta espécie predominou em quatro parcelas. Na parcela 3 o número de indivíduos de Laguncularia racemosa foi igual a Rhizophora mangle e na parcela 5 Rhizophora mangle apresentou maior número de indivíduos. Com relação ao número de troncos, Rhizophora mangle apresenta menor número de ramificações do que as duas outras espécies. O maior número de troncos foi observado na parcela 6, onde as árvores de Laguncularia racemosa e Avicennia schaueriana apresentaram-se bem ramificadas, demonstrando a ocorrência de um estresse gerado por corte de madeira, fato que foi corroborado pelas observações realizadas no campo nos trechos de manguezal próximos a esta parcela.
A maior contribuição para a área basal total em todas parcelas foi da espécie Avicennia schaueriana em função da presença de exemplares de grande porte neste manguezal, principalmente nas parcelas 2, 3 e 4, fato que pode ser confirmado através da observação dos valores de área basal desta espécie nestas parcelas. Laguncularia racemosa foi a espécie que mais contribui para a área basal nas parcelas 1 e 6, enquanto que Rhizophora mangle apresentou maior contribuição na parcela 5.
Neste perfil a espécie Laguncularia racemosa apresentou maior destaque com valores superiores de número de indivíduos (Ind.), densidade relativa (DR), dominância relativa (DoR), índice de valor de importância (IVI) e índice de valor de cobertura (IVC) do que as outras duas espécies. A espécie Avicennia schaueriana apresentou maior contribuição para área basal.
Neste bosque a fisiografia é de manguezal de franja. Os resultados obtidos para análise fitossociológica corroboraram a avaliação da estrutura deste perfil.
A análise fitossociológica do bosque do perfil do manguezal da Pousada do Bosque está apresentada na Tabela 32.
Tabela 32 - Avaliação fitossociológica do bosque no perfil do manguezal da Pousada do Bosque.
Pousada do Bosque Parcela |
Ind |
Troncos |
Área basal |
| Rm |
Lr |
As |
Rm |
Lr |
As |
Rm |
Lr |
As |
| 1 |
0 |
37 |
0 |
0 |
62 |
0 |
0 |
0,242351 |
0 |
| 2 |
0 |
29 |
0 |
0 |
51 |
0 |
0 |
0,302237 |
0 |
| 3 |
0 |
49 |
0 |
0 |
77 |
0 |
0 |
0,282968 |
0 |
| Soma |
0 |
115 |
0 |
0 |
190 |
0 |
0 |
0,827556 |
0 |
| Total
|
|
115 |
|
|
190
|
|
|
0,827556
|
|
| Espécie |
Parcelas |
Ind. |
AB |
DR |
FA |
FR |
DoR |
IVI |
IVC |
| R. mangle |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0,00 |
0 |
| L.racemosa |
3 |
115 |
0,82 |
1 |
100 |
100 |
1 |
102,00 |
2 |
| A schaueriana |
0 |
0 |
0 |
0 |
0 |
0 |
0 |
0,00 |
0 |
| |
3 |
115 |
0,82 |
1 |
100 |
100 |
1 |
|
|
Rm - Rhizophora mangle; Lr - Laguncularia racemosa; As - Avicennia schaueriana; AB - Área basal; DR - Densidade relativa; FA - Freqüência absoluta; FR - Freqüência relativa; DoR - Dominância relativa; IVI - Índice de valor de importância; IVC - Índice de valor de cobertura.
Os dados da Tabela 32 indicam a ocorrência monoespecífica de Laguncularia racemosa nos perfis. Todas as parcelas apresentam um elevado número de troncos que pode ser oriundo do estresse por deposição de areia a que o bosque está submetido. A maior contribuição para a área basal ocorreu na parcela 2. Este bosque apresenta fisiografia ribeirinha.
Considerações Finais
A espécie Rhizophora mangle apresentou maior abundância nos perfis da praia do Recife e da ilha do Jorge, enquanto o perfil do Bracuí apresentou maior abundância de Laguncularia racemosa. Isto permitiu caracterizar os manguezais do Bracuí e Pousada do Bosque como bosques brancos e os manguezais da praia do Recife e da ilha do Jorge como bosques vermelhos.
Rhizophora mangle e Laguncularia racemosa apresentaram os maiores números totais de exemplares nos perfis estudados. Com relação ao percentual de ocorrência, tanto Rhizophora mangle quanto Laguncularia racemosa apresentaram participações de próximas a 46%, enquanto Avicennia schaueriana apenas9%.
A maior densidade de exemplares ocorreu no perfil do manguezal da praia do Recife onde foram estimados 44.000 ind/ha. A menor densidade estimada foi a do perfil do manguezal do Bracuí com 23.625 ind/ha.
Os maiores valores médios de DAP e altura foram observados no perfil do manguezal da Ilha do Jorge. O menor valor de DAP médio foi registrado no perfil do manguezal do Bracuí e o menor valor para altura média foi mensurado no perfil do manguezal da praia do Recife.
A maior contribuição para área basal do manguezal do Bracuí foi a da classe diamétrica do DAP ³ 5 e < 10. A maior contribuição para a área basal tanto do manguezal da praia do Recife quanto da Ilha do Jorge foi da classe diamétrica do DAP ³ 10. Isto permitiu concluir que no Bracuí o manguezal encontra-se em estado de regeneração avançada e que os manguezais da praia do Recife e da Ilha do Jorge encontram-se mais maduros estruturalmente.
A espécie que mais contribuiu para a área basal total dos cinco perfis foi Rhizophora mangle.
A análise fitossociológica indicou a ocorrência de um grande número de exemplares nas parcelas estudadas. O grande número de troncos foi registrado sempre nas Parcelas iniciais dos perfis, característica oriunda do estresse causado pelo desmatamento que ocorre preferencialmente nas regiões do manguezal de acesso mais fácil, próximas a água, para facilitar o transporte dos troncos após o corte. Este processo também é muito observado nos manguezais da baía de Guanabara e baía de Sepetiba.
No perfil do Bracuí a espécie Laguncularia racemosa foi a mais importante e apresentou maiores valores para número de indivíduos (Ind.), densidade relativa (DR), freqüência absoluta (FA), freqüência relativa (FR) índice de valor de importância (IVI) e índice de valor de cobertura (IVC).
No perfil da praia do Recife e da ilha do Jorge a espécie Rhizophora mangle foi a mais importante e apresentou valores superiores de número de indivíduos (Ind.), área basal (AB), densidade relativa (DR), freqüência relativa (FR), dominância relativa (DoR), índice de valor de importância (IVI) e índice de valor de cobertura (IVC). Nos perfis do Ariró e da Pousada do bosque, os dados permitiram concluir que a espécie mais importante foi Laguncularia racemosa.
As fisiografias dos manguezais estudados são de franja (Bracuí, praia do Recife, ilha do Jorge Grego e do rio Ariró) e ribeirinho (Pousada do Bosque).
A fauna observada nos manguezais dos cinco perfis foi representada por moluscos e crustáceos. A malacofauna foi representada por Anomalocardia brasiliana (berbigão), Lucina pectinata (ameixa), Iphigenia brasiliensis (tarioba), Teredo spp. (broca da madeira) e Ostrea rhizophorae (ostra do mangue), Mytella spp.(sururu), Tagelus plebeius (unha-de-velho), Neritina virginica e Littorina (Scabra) angulifera (buso do mangue). Salienta-se que estes animais são fundamentais para a subsistência das populações pesqueiras locais. A presença de bancos de Mytella spp., como os observados no manguezal da ilha do Jorge, são raros nos manguezais do estado do Rio de Janeiro. Sendo assim sugere-se que sejam implementados estudos destas populações e sejam elaborados mecanismos para sua preservação.
A carcinofauna foi representada por Chthamalus spp. (craca), Balanus spp. (craca), Callinectes spp. (siri), Uca (Minuca) rapax (chama-maré), Uca (Celuca) leptodactyla (chama-maré), Aratus pisonii (marinheiro), Goniopsis cruentata (aratu vermelho e preto), Chasmagnatus granulata (catanhém), Ucides cordatus (caranguejo uçá) e Cardisoma guanhumi (guaiamum). Em todos os manguezais estudados foi observada a intensa captura de exemplares de Ucides cordatus e Cardisoma guanhumi através de técnicas predatórias. A presença de caranguejeiros do Rio de Janeiro atuando nos manguezais de Angra dos Reis foi confirmada com auxílio de pescadores locais e da observação de acampamento no manguezal. Este fenômeno migratório para captura do caranguejo já havia sido constado através dos estudos efetuados por Alves (comunicação pessoal) nos manguezais da Baía de Guanabara. Caso este fenômeno se mantenha os estoques de caranguejo dos manguezais de Angra dos Reis ficarão ameaçados.
Nos manguezais estudados foram evidenciados impactos como desmatamento, aterros, construção de residências, construção de estradas, alteração do padrão de circulação das águas e lançamento de lixo. Estes impactos são frutos do processo de ocupação desordenada que, na região sul-fluminense, são decisivos para a redução das áreas de manguezais. Outro ponto a destacar é a ausência de estudos aprofundados destes ecossistemas na região. É fundamental conhecer bem estes ambientes para que sejam tomadas decisões e sejam implementadas medidas para controlar o grau de degradação atual dos manguezais da região.
A ocorrência de uma grande número de troncos observado em parcelas do Manguezal do Ariró (parcela 6) e da Pousada do Bosque (todas) são reflexos do corte de madeira e da deposição de areia respectivamente. Apesar de apresentarem boa estrutura de bosque em todos os manguezais estudados foi observado um intenso corte de troncos de mangue.
Assim, baseando-se nas informações obtidas para elaboração deste relatório, pode-se listar os manguezais estudados de acordo com seu estado de conservação em: Ariró; praia do Recife; Pousada do Bosque; ilha do Jorge e Bracuí.
7.2. Flora Aquática - topo
7.2.1. Hidroquímica - topo
É feito todo um acompanhamento periódico dos aspectos físico-químicos das águas de Itaorna e do Saco Piraquara de Fora. A Eletronuclear se encarrega de realizar este monitoramento nos seguintes pontos: Z3 que se encontra em Itaorna, além de Z4 e 47B no Saco Piraquara de Fora. Para isto, utiliza-se dos seguintes parâmetros: oxigênio dissolvido, nitrito, nitrato, silicato e fosfato; a freqüência de amostragem é mensal, mas sua finalização é através de médias anuais para o período de 1991 a 2000.
As concentrações de oxigênio dissolvido variaram nos cincos primeiros anos de análise de 4,62 mL.L-1 na estação Z4 e 4,82 mL.L-1 tanto para Z3 quanto para 47B, havendo um excelente índice de concentração de oxigênio entre estes pontos, já que encontram-se sempre acima de 4,20 mL.L-1, que é o limite para a Classe 5 da Resolução Conama nº 20/86. Esta Classe é a de águas salinas destinadas à recreação de contato primário, proteção das comunidades aquáticas, e criação natural e/ou intensiva (aquicultura) de espécies destinadas à alimentação humana. No ano de 1997 (Tenenbaum et al., 1997), houve uma variação quanto a essa concentração de 2,42 mL.L-1 em Z4 pelo mês de dezembro a 6,66 mL.L-1 em Z3 no mês de abril, sendo observadas concentrações mais altas durante o verão (de janeiro a abril), havendo uma tendência análoga, com exceção de Z3 nos meses de julho, novembro e dezembro. Em 1998 (Tenenbaum, 1999) houve um aumento dessa concentração em ambos os pontos, pois chegou a 3,17 mL.L-1 em Z3 no mês de janeiro, a 8,54 mL.L-1 em Z3 no mês de março, e de um modo geral, os três pontos de coleta apresentaram o mesmo padrão de variação.
No ano de 1999 esse parâmetro variou de 3,32 mL.L-1 (Z4 em abril) a 5,50 mL.L-1 (Z4 em novembro) e as concentrações mais altas (próximas de 5,0 mL.L-1) (Tenenbaum et al, 2000) deste ano de estudo foram observadas nos meses da primavera, comparando-se com a Classe 5 da Resolução Conama nº 20/86, houve uma redução na concentração, que poderia de certa forma ser prejudicial, mas neste caso não chega a ser tão agravante para a região. Já em 2000, houve um relativo aumento nessa concentração, sendo de 4,10 mL.L-1 em Z4 no mês de janeiro a 7,14 mL.L-1 no ponto 47B em agosto (Tenenbaum et al, 2001). Este aumento deu-se do início para o final do ano, destacando-se ainda um aumento nos valores de Z4 entre os meses de abril e julho.
Os valores de nitrato e nitrito encontram-se sempre abaixo do limite para a Classe 5, que é de 0,07 e 0,71 µM para nitrito e nitrato, respectivamente. Isso foi facilmente observado nos cinco primeiros anos de análise, onde as concentrações de nitrito e nitrato foram respectivamente de 0,02 µM e 0,11 µM em Z3, 0,02 µM e 0,13 µM em Z4 e 0,02 µM e 0,12 µM em 47B. Em 1997, os teores de nitrito variaram de não detectável a 0,12 µM, enquanto que os teores de nitrato foram de 0,02 µM em Z3 no mês de dezembro a 1,65 µM em Z3 no mês de agosto, havendo uma oscilação nos níveis de medição em torno do valor médio de 0,40 µM (Tenenbaum et al, 1997), com uma tendência análoga nas três estações de coleta. Já em 1998, os números foram mantidos em relação ao ano anterior quanto os teores de nitrito, havendo, quanto aos teores de nitrato, uma variação de 0,02 µM em Z4 no mês de janeiro a 2,73 µM no ponto 47B em março (Tenenbaum & Villac, 1999), mantendo-se o mesmo padrão de variação nos três pontos de coleta, com exceção nos meses de março e junho deste mesmo ano.
Ainda quanto às medidas de nitrito em 1999, observou-se uma variação de não detectável a 0,31 µM e de nitrato, uma variação de 0,02 µM em 47B no mês de dezembro a 0,87 µM em Z4 no mês de julho (Tenebaum et al, 2000), mantendo-se valores bem próximos entre si nos três pontos, com exceção nos meses de julho e agosto. Mas em 2000, os valores subiram, de nitrito indo de não detectável a 0,41 µM no ponto 47B em julho e nitrato variando de 0,01 µM em 47B no mês de abril a 9,50 µM em Z4 no mês de junho (Tenenbaum et al, 2001), mas como este último valor foi absurdamente alto conclui-se que pode ter havido um erro de análise, sendo assim desconsiderado.
Os níveis de fosfato nas três estações encontraram-se sempre bem próximos, sendo a média igual à 0,01 µM. E o valor máximo de fosfato detectado nos primeiros cinco anos de medição foi de 0,03 µM, mas neste caso a Resolução Conama nº 20/86 não prevê limites para este parâmetro, como foi mencionado em oxigênio dissolvido, nitrito e nitrato. Nos anos seguintes como em 1997 houve uma variação média de 0,03 µM em Z3 no mês de novembro a 0,51 µM em Z4 no mês de agosto (Tenenbaum et al, 1997), mantendo-se praticamente estável em todos os meses do ano com uma diferença apenas entre janeiro e março. Em 1998, essa variação foi de 0,02 µM no ponto Z3 em março a 0,64 µM em 47B no mês de março (Tenenbaum & Villac, 1999). A variação de 1999, manteve-se no mesmo padrão dos últimos anos, entre 0,01 µM em Z3 no mês de dezembro a 0,62 µM em Z4 no mês de junho (Tenenbaum et al, 2000), observando-se níveis mais baixos de fosfato no início do ano (entre janeiro e maio) e em dezembro. E em 2000 esse parâmetro oscilou entre 0,01 µM em agosto e 0,54 µM no ponto Z4 em junho, havendo uma elevação desses valores entre maio-junho (Z3 com valores mais baixos) e setembro (Tenenbaum et al, 2001).
Os valores de silicato encontram-se sempre abaixo de 0,32 µM, tendo, dentro dos cinco primeiros anos, uma média anual de 0,15 µM para a estação Z3, em Z4 e 47B valores foram de 0,17 e 0,16 µM, respectivamente, não havendo também medidas de padrão previstas para este parâmetro pela Resolução Conama nº 20/86. No ano de 1997, os valores foram de 0,31 µM (Z3, junho) a 11,04 µM (Z4, setembro) (Tenenbaum et al, 1997), observando-se um aumento paulatino de julho a agosto, com máxima em setembro e uma queda brusca entre outubro e novembro, e um novo aumento em dezembro. Em 1998, houve variação de 0,60 µM em Z4 no mês de março a 11,58 µM na estação Z4 em junho (Tenenbaum & Villac, 1999), com um mesmo padrão nas três estações de coleta. Os teores de silicato, em 1999, variaram de 0,75 µM em Z3 no mês de dezembro a 9,51 µM no ponto Z4 pelo mês de agosto (Tenenbaum et al, 2000), sendo que este mesmo ponto apresentou teores de silicato mais altos que os outros dois pontos de coleta ao longo de todo o ano. Em 2000, o ponto Z4 ainda manteve o nível de silicato muito alto em relação aos demais pelo menos até o mês de agosto, mas os valores em si variaram de 0,28 µM em 47B no mês de dezembro a 6,97 µM na estação Z4 em julho (Tenenbaum et al, 2001), tendo havido um declínio do valor máximo em relação aos demais anos de análise.
Segundo o estudo realizado por Villac (1983), os maiores valores de clorofila a foram observados no verão e os menores no inverno, estando esta variação influenciada principalmente pela precipitação. Foi necessário então fazer um acompanhamento dos impactos do efluente térmico da usina (Angra 1, que foi ativada em dezembro de 1981, iniciando suas operações em 1985) sobre o fitoplâncton da região. Desta forma, o Instituto de Biologia da Universidade Federal do Rio de Janeiro produziu relatórios de monitoramento da região e os dados foram fornecidos por Furnas Centrais Elétricas , referentes a esses relatórios de monitoramento produzidos no período de 1987 a 2000 (Tenenbaum & Villac, 1990; Tenenbaum & Nascimento, 1993; Tenenbaum & Nascimento, 1994; Tenenbaum & Dias, 1995; Tenenbaum & Villac, 1996; Tenenbaum et al, 1997; Tenenbaum & Villac, 1999; Tenenbaum et al, 2000 e Tenenbaum et al, 2001).
Neste monitoramento são coletadas mensalmente amostras a 5 m de profundidade em 3 pontos fixos: o ponto Z3 que está localizado na enseada de Itaorna, onde ocorre captação da água para resfriamento do reator. Os pontos Z4 e 47B localizam-se no saco Piraquara de Fora, onde é feito o lançamento do efluente térmico da usina, foram analisados os dados de clorofila a e as correlações com variáveis abióticas (temperatura, salinidade, transparência da água, pluviosidade e nutrientes inorgânicos. No monitoramento referente ao período de 1987 a 1989 (Tenenbaum & Villac, 1990), os valores de clorofila a giraram em torno de 0,81 µg.L-1 podendo ser considerado um ecossistema oligotrófico (Sassi & Kutner, 1982). No período de 1989 a 1991 Tenenbaum et al (1991), registrou-se baixa concentração de clorofila a numa média de 1,31 µg.L-1, talvez indicando uma sazonalidade para o fitoplâncton influenciada principalmente pelo regime de chuvas.
Entre janeiro e dezembro de 1993, o ambiente foi considerado oligotrófico, com uma média de 0,75 µg.L-1 para clorofila a que foi considerada bastante baixa em relação aos estudos de monitoramento anteriores. Já o monitoramento de 1994 (Tenenbaum & Dias, 1995), registrou uma média também baixa de clorofila, girando em torno de 0,77 µg.L-1, assim como em estudos anteriores, caracterizando o ambiente como oligotrófico. Os monitoramentos, a partir de 1997, registraram médias maiores de clorofila a, com exceção do ano de 2000, em que ocorre uma diminuição dos valores.
Em 1997, a variação foi de 0,27 µg.L-1 no ponto 47B no mês de agosto a 2,61 µg.l-1 em Z3 no mês de abril, em que as concentrações mais altas foram observadas no verão (fevereiro - abril) e em novembro (Tenenbaum et al, 1997). No ano de 1998 (Tenenbaum & Villac, 1999), os valores ficaram entre 0,23 µg.L-1 no ponto 47B em agosto e 4,03 µg.L-1 também na 47B só que em março, com uma variação análoga nas três estações. Durante 1999 (Tenenbaum et al, 2000), a estação Z4 apresentou valores mais altos que os demais em vários meses do ano, mas os valores oscilaram bastante entre 0,25 µg.L-1 no 47B em janeiro e 3,28 µg.L-1 em Z4 no mês de dezembro (que foi o maior registro do ano). Já em 2000, a variação foi de 0,31 µg.L-1 em 47B no mês de agosto a 1,64 µg.L-1 na estação Z3 em janeiro, apesar de ter havido uma mesma tendência para os três pontos de coleta durante o período de estudo, observou-se uma divergência entre os pontos, especialmente em janeiro, julho e novembro, sendo que não foram executadas análises em maio em função de detritos (Tenenbaum et al, 2001).
Assim, a comunidade fitoplanctônica da região é influenciada principalmente pelas variações sazonais, observadas através das oscilações dos fatores meteorológicos e hidrográficos.
7.2.1.1. Caracterização da Comunidade Planctônica - topo
A caracterização da comunidade planctônica foi feita através das análises dos parâmetros físicos, químicos, clorofila a e do fitoplâncton, zooplâncton e ictioplâncton, com base em amostras coletadas em Itaorna e no Saco Piraquara de Fora, com a distribuição das estações de coleta em radiais, conforme mostra a Figura 27, em um ciclo nietermal de 48 horas, em duas estações, com coletas de 6 em 6 horas em marés de enchente e vazante.
Em Itaorna a coleta foi feita no ponto Z3R, próximo ao local previsto para a captação de água para o resfriamento dos condensadores dos sistema de água de circulação de Angra 3, entre a Praia de Fora do Mamede e a Ilha Comprida, na direção da Ponta do Coqueirão, e, a partir deste ponto, nas radiais R5, R6, R7, R8 e R9. No Saco Piraquara de Fora, a coleta já foi feita nos pontos Z4R, próximo ao local previsto para a saída da água de resfriamento (água captada misturada com água doce e cloro), e 47BR, bem como nas radiais R1 e R2, situadas entre os pontos Z4R e 47BR, e R3 e R4, situadas após o ponto 47BR. Os pontos Z3, Z4 e 47BR foram estabelecidos pela Eletronuclear, sendo os demais marcados para permitir traçar um melhor perfil nas áreas de influência direta e indireta da usina.
Em uma primeira etapa, a coleta foi feita em todas as radiais. No ciclo de 24 horas, foram coletadas amostras apenas nos pontos Z3 e Z4, por serem monitorados pela Eletronuclear.
As amostras foram coletadas com garrafa de Niskin de 10 L (General Oceanics), revestida com teflon, em cada profundidade amostrada. A primeira subamostra drenada da garrafa de coleta, destinada à determinação do teor de oxigênio dissolvido, foi colocada em um frasco de DBO de 300 mL e imediatamente medida com o emprego de oxímetro Orion (Model 835). O pH foi também determinado imediatamente (15 minutos no máximo), por meio de potenciômetro Corning (Digital pH-10 Chekmite), e calibrado com tampões de pH 7,0 e 10,0, conforme descrito por Grasshoff et al (1983). Em seguida foi medida a salinidade, com o emprego do condutivímetro Handylab LF 1 (Schot) adquadamente calibrado.
Figura 27 - Localização das estações utilizadas para as coletas de parâmetros físicos, químicos, clorofila a, fitoplâncton, zooplâncton e ictioplâncton.
A segunda subamostra drenada da garrafa, destinada à determinação de nutrientes, foi colocada em um frasco de polipropileno de 1000 mL, previamente lavado com HCl 1:1 e com água ultrapura (Milli-Q Gradient UV), e congelada em freezer.
Para a determinação do material particulado em suspensão, a subamostra foi drenada da garrafa para um frasco de polipropileno de 2000 mL e rapidamente (1 hora no máximo) filtrada a vácuo, sob pressão máxima de 250 mmHg, em filtros de fibra de vidro Millipore AP15, previamente identificados, pesados em laboratório e acondicionados em placas de Petri individuais. Uma vez utilizados, os filtros foram guardados em suas respectivas placas de Petri e armazenados em saco plástico devidamente identificado.
Para a determinação de clorofila a, uma subamostra foi coletada em frasco plástico verde de 2000 mL e imediatamente (15 minutos no máximo) filtrada a vácuo, sob pressão máxima de 250 mmHg, em membranas de celulose Millipore HAWP com poros de 0,45 µm. Durante o processo de coleta e filtração, as amostras foram protegidas da luz. Os filtros foram colocados em tubos criovials e em seguida congelados e armazenados em galão de nitrogênio líquido.
Em laboratório, as amostras foram analisadas em triplicata para cada parâmetro. O ortofosfato reativo foi determinado pelo método fosfomolibídico (Grasshoff et al., 1983), o mesmo empregado para fósforo total, sendo que, neste caso, as amostras foram digeridas em meio ácido com persulfato de potássio. O nitrogênio amoniacal (N-NH3+N-NH4+), ou simplesmente amônia, foi determinado pelo método do azul de indofenol (Parsons et al., 1984). O nitrito foi determinado pelo método da diazotação (Grasshoff et al., 1983) e o nitrato, por redução em coluna de Cd-Cu seguido de diazotação (Grasshoff et al., 1983), tendo sido analisado po meio de injeção em fluxo. O silicato foi determinado pelo método silicomolibídico (Grasshoff et al., 1983).
O limite de detecção dos métodos foi estimado como sendo três vezes o desvio padrão dos brancos, multiplicado pelo fator de calibração. Não se dispôs de padrão certificado para a determinação de amônia e o limite de detecção para este ensaio foi estimado em 0,05 µM. Os limites de detecção adotados para a determinação de ortofosfato, nitrito, nitrato e silicato, obtidos mediante determinação de soluções-padrão OSI, estão apresentados na Tabela 33.
Tabela 33 - Limites de detecção adotados para determinação de ortofosfato, nitrito, nitrato e silicato
| Parâmetro
|
Limite de detecção |
| P-PO43-
|
0,02 µM |
| N-NO2-
|
0,02 µM |
| N-NO3-
|
0,05 µM |
| Si-SiO2-
|
0,02 µM |
A concentração de material particulado em suspensão (MPS) foi determinada gravimetricamente em filtros Millipore AP15, previamente calcinados a 450ºC por 3 horas e pesados no laboratório em uma balança analítica com precisão de x mg. Para cada amostra foram filtrados de 1 a 2 litros, sendo os filtros secos em estufa a 40ºC até peso constante e repesados.
A clorofila a foi determinada após extração em acetona a 90%, por 18 horas, a 4°C. As medições no fluorímetro Turner Designs TD-700 e os cálculos seguiram os procedimentos e as equações descritas por Parsons et al. (1984), com enfoque de metrologia (Mattos, 2001; Paiva, 2001).
Nas duas opções de detecção o aparelho foi calibrado com clorofila a pura (Sigma C-6144) e o limite de detecção foi de 0,02 µg.L-1 (Mattos, 2001).
Os parâmetros foram analisados na superfície e a 5 m de profundidade nos pontos Z3R a R9, em Itaorna, e Z4R a R4, do Saco Piraquara de Fora. Todos os dados obtidos na campanha para avaliar a distribuição espacial estão dispostos na Tabela 34 e na Tabela 35.
Na região estudada, os valores de temperatura oscilaram entre 24,2 e 28,7ºC, sendo a média de 25,6º C. Desta forma, apresentaram baixa variabilidade (CV 5%), sendo que o maior valor foi registrado em Itaorna (radial de Z3 a R9) e o menor, no Saco Piraquara de Fora (radial de Z4R a R4), ambos na superfície.
Em quatro dos seis pontos de Itaorna, as temperatura na camada de fundo (5 m) foram levemente superiores às da superfície. Já nas estações do Saco Piraquara de Fora, foi possível observar uma tênue estratificação, com o menor valor sendo encontrado a 5 m de profundidade (Figura 28)
Os valores de salinidade também apresentaram baixa variabilidade (CV 2%) e a camada de fundo mostrou-se mais homogênea que a de superfície (Figura 29). O maior valor (32,9 S) foi registrado no Saco Piraquara de Fora, na superfície da estação 47BR, e o menor (30,8 S), em Itaorna, na estação R7, também na superfície.
Tabela 34 - Dados coletados em maio de 2002 para avaliar a distribuição espacial, em Itaorna e no Saco Piraquara de Fora - temperatura, salinidade, oxigênio, pH, ortofosfato e fósforo total
Estações |
Horário de coleta |
Temp.
(ºC)
|
Salinidade
(S)
|
Oxigênio
(mL.L-1)
|
pH
|
Ortofosfato
(µM P-PO4-3)
|
Fósforo total
(µM P) |
| sup |
5 m |
sup |
5 m |
sup |
5 m |
sup |
5 m |
sup |
5 m |
sup |
5 m |
| Z4 R |
14:55 |
28,6 |
25,6 |
32,4 |
32,6 |
5,11 |
4,48 |
8,00 |
8,00 |
0,20 |
0,23 |
1,32 |
0,87 |
| R1 |
15:35 |
27,5 |
25,6 |
32,7 |
32,7 |
4,76 |
4,41 |
8,08 |
8,12 |
0,19 |
0,21 |
0,37 |
0,37 |
| R2 |
16:05 |
28,7 |
25,4 |
32,5 |
32,8 |
4,83 |
4,61 |
8,00 |
8,21 |
0,19 |
0,21 |
0,40 |
0,49 |
| 47 BR |
16:50 |
26,8 |
27,1 |
32,9 |
32,7 |
4,55 |
5,11 |
8,14 |
8,07 |
0,17 |
0,15 |
0,28 |
0,53 |
| R3 |
17:10 |
26,3 |
24,9 |
32,8 |
32,8 |
4,61 |
4,41 |
8,21 |
8,13 |
0,17 |
0,17 |
0,35 |
0,29 |
| R4 |
17:50 |
26,4 |
25,0 |
32,6 |
32,7 |
5,11 |
4,83 |
8,25 |
8,28 |
0,15 |
0,18 |
0,29 |
0,28 |
| Z3 R |
10:30 |
24,6 |
24,7 |
31,9 |
32,2 |
3,78 |
4,62 |
8,27 |
8,28 |
0,14 |
0,13 |
0,41 |
0,57 |
| R5 |
11:45 |
24,2 |
24,3 |
31,7 |
32,3 |
5,11 |
5,18 |
8,21 |
8,19 |
0,19 |
0,16 |
0,66 |
0,43 |
| R6 |
12:15 |
24,5 |
24,6 |
31,3 |
32,0 |
4,97 |
4,83 |
8,21 |
8,14 |
0,16 |
0,14 |
0,52 |
0,24 |
| R7 |
13:15 |
24,2 |
24,5 |
30,8 |
32,5 |
4,83 |
4,62 |
8,12 |
8,21 |
0,18 |
0,19 |
0,47 |
0,44 |
| R8 |
13:40 |
25,3 |
25,0 |
32,0 |
32,3 |
4,83 |
4,97 |
8,08 |
8,13 |
0,13 |
0,15 |
0,72 |
0,37 |
| R9 |
14:20 |
25,7 |
25,3 |
31,7 |
32,5 |
4,97 |
4,90 |
8,12 |
8,00 |
0,07 |
0,09 |
0,58 |
0,55 |
Tabela 35 - Dados coletados em maio de 2002 para avaliar a distribuição espacial, em Itaorna e no Saco Piraquara de Fora - amônia, nitrito, nitrato, silicato, MPS e clorofila a.
Estações |
Horário de coleta |
Amônia
(µM NH3) |
Nitrito
(µM N-NO2-) |
Nitrato
(µM N-NO3-) |
Silicato
(µM Si-SiO2) |
MPS
(mg.L-1)
|
Clorofila a
(µg.L-1) |
| sup |
5 m |
sup |
5 m |
sup |
5 m |
sup |
5 m |
sup |
5 m |
sup |
5 m |
| Z4 R |
14:55 |
0,05 |
0,95 |
0,08 |
0,20 |
0,67 |
0,12 |
4,74 |
6,40 |
30,20 |
31,80 |
0,529 |
4,58 |
| R1 |
15:35 |
0,05 |
0,05 |
0,03 |
0,05 |
0,05 |
0,05 |
3,93 |
2,97 |
32,10 |
33,00 |
2,258 |
6,17 |
| R2 |
16:05 |
0,07 |
0,05 |
0,05 |
0,04 |
0,05 |
0,05 |
3,39 |
3,33 |
32,40 |
18,00 |
1,886 |
4,86 |
| 47 BR |
16:50 |
0,05 |
0,11 |
0,02 |
0,02 |
0,16 |
0,05 |
3,94 |
4,41 |
30,70 |
29,90 |
2,267 |
3,19 |
| R3 |
17:10 |
0,05 |
0,05 |
0,04 |
0,05 |
0,05 |
0,05 |
2,93 |
3,34 |
37,90 |
46,20 |
2,224 |
0,21 |
| R4 |
17:50 |
0,05 |
0,05 |
0,02 |
0,02 |
0,05 |
0,05 |
3,59 |
3,67 |
35,70 |
43,80 |
2,195 |
1,87 |
| Z3 R |
10:30 |
0,05 |
0,05 |
0,14 |
0,13 |
0,17 |
0,05 |
4,43 |
3,13 |
14,85 |
27,20 |
0,351 |
1,02 |
| R5 |
11:45 |
0,05 |
0,05 |
0,02 |
0,02 |
0,68 |
0,56 |
7,53 |
5,15 |
46,57 |
33,50 |
3,353 |
2,63 |
| R6 |
12:15 |
0,05 |
0,05 |
0,03 |
0,04 |
0,05 |
0,05 |
6,78 |
4,18 |
27,20 |
17,35 |
3,733 |
2,72 |
| R7 |
13:15 |
0,05 |
0,05 |
0,04 |
0,03 |
0,05 |
0,05 |
10,62 |
3,57 |
32,37 |
29,60 |
4,909 |
1,95 |
| R8 |
13:40 |
0,07 |
0,05 |
0,03 |
0,06 |
0,05 |
0,36 |
5,46 |
3,82 |
29,30 |
32,00 |
3,451 |
2,46 |
| R9 |
14:20 |
0,05 |
0,05 |
0,07 |
0,09 |
0,05 |
0,56 |
6,60 |
5,48 |
29,10 |
30,90 |
5,073 |
2,23 |
Figura 28 - Distribuição espacial da temperatura (ºC)
Figura 29 - Distribuição espacial da salinidade (S)
Como observado para temperatura, os valores de salinidade encontrados nas estações de Itaorna foram ligeiramente mais elevados que os valores de superfície. Já nas estações do Saco Piraquara de Fora, os valores foram mais homogêneos ao longo da coluna d’água. Tendo em vista esses valores de salinidade a Resolução Conama nº 20/86 classifica essas águas como salinas, por apresentarem valores iguais ou superiores a de 30 S.
A distribuição do oxigênio dissolvido mostrou-se homogênea nas duas radiais (Figura 30) apresentando baixa variabilidade entre os dados (CV 7%). O maior (5,18 mL.L-1) e o menor valor (3,78 mL.L-1) foram registrados, respectivamente, na estação R5, no fundo (5 m), e na estação Z3R, na superfície, ambas em Itaorna. A menor média de oxigênio dissolvido encontrada foi de 4,20 mL.L-1, que é o valor mínimo estabelecido pela Resolução Conama nº 20/86 para águas salinas da classe 5 (classe em que se enquadra a área em questão). Essas águas são destinadas a recreação de contato primário, proteção das comunidades aquáticas e criação natural e/ou intensiva (aqüicultura) de espécies destinadas à alimentação humana.
Figura 30 - Distribuição espacial de oxigênio de dissolvido (mL.L-1)
As águas da região estudada apresentaram grande homogeneidade em relação aos valores de pH, que apresentaram a menor variabilidade observada entre os dados (CV 1%). Os valores oscilaram entre 8,00 e 8,28 e ficaram muito próximos, tanto na superfície quanto a 5 m de profundidade. Levando em conta a variação espacial, homegeneidade desse parâmetro é grande, tanto em Itaorna quanto no Saco Piraquara de Fora (Figura 31).
Figura 31 - Distribuição espacial do pH
Para o ortofosfato foram registrados valores oscilando entre 0,07 e 0,23 µM, apresentando assim moderada variabilidade (CV 23%). O valor mais elevado foi encontrado a 5 m de profundidade, no Saco Piraquara de Fora, na estação Z4R, e os dois menores, em Itaorna, na superfície (0,07 µM) e no fundo (0,09 µM), da estação R9 (Figura 32).
Figura 32 - Distribuição espacial do ortofosfato (µM)
O fósforo total apresentou uma considerável variabilidade entre os dados (CV 47%), que oscilaram entre 0,24 e 1,32 µM. O maior e o menor valor foram registrados, respectivamente, na superfície (# Z4-R) e no fundo (# R6). Nas estações de Itaorna, os valores de fósforo total foram levemente superiores na superfície, o que pode indicar aporte desse nutriente proveniente do ambiente terrestre. Como citado anteriormente, o maior valor desse parâmetro foi registrado na estação Z4R, que é a mais próxima da costa no Saco Piraquara de Fora, que recebe o lançamento de água proveniente das usinas (Figura 33).
Figura 33 - Distribuição espacial do fósforo total (µM)
A amônia apresentou a maior variabilidade entre todos os parâmetros analisados (CV 285%), pois seus valores oscilaram entre o limite de detecção do método (<0,05 µM) e 0,95 µM, registrado na estação Z4R, a 5 m de profundidade. Os maiores valores desse parâmetro foram registrados nas estações do Saco Piraquara de Fora a 5 m de profundidade (Figura 34).
O nitrito apresentou valores entre não detectável (<0,02 µM) e 0,20 µM, com uma considerável variabilidade (CV 87%). Os valores mais elevados foram registrados nas estações Z4R (0,20 µM), no Saco Piraquara de Fora, e Z3R (0,13 µM), em Itaorna, em ambos os casos no fundo, podendo ser notado que esse nutriente apresenta uma distribuição espacial com os maiores valores na camada do fundo e nas estações mais próximas à costa (Figura 35).
Figura 34 - Distribuição espacial de amônia (µM)
Figura 35 - Distribuição espacial de nitrito (µM)
O nitrato apresentou uma distribuição espacial semelhante à do nitrito, com os valores mais elevados na camada do fundo e nas estações mais próximas à costa (Figura 36), tendo sido registrados valores desde não detectável (<0,05 µM) até 0,68 µM e, conseqüentemente, uma alta variabilidade (CV 173%). Os dois valores mais elevados, 0,68 e 0,67 µM, ocorreram na superfície, em Itaorna (# R5) e no Saco Piraquara de Fora (# Z4R), respectivamente. Apesar desses resultados, a maior parte das estações apresentou valores de nitrato abaixo do nível de detecção do método.
O maior valor de silicato (10,62 µM) foi registrado em Itaorna (# R7) e o menor (2,93 µM), no Saco Piraquara de Fora, ambos na superfície. Este parâmetro apresentou uma considerável variabilidade entre os dados (38%). Nas estações de Itaorna (Z3 a R9), a concentração de silicato foi maior na superfície. Já nas estações do Saco Piraquara de Fora (Z4R a R4), esse padrão se repetiu apenas em dois pontos (R1 e R2) (Figura 37). Esses valores estão de acordo com os indicados em estudos anteriores realizados na região (Tenenbaum et al. 2000).
Figura 36 - Distribuição espacial de nitrato (µM)
Figura 37 - Distribuição espacial de silicato (µM)
Os valores de material particulado em suspensão (MPS) não obedeceram a qualquer padrão de distribuição espacial, na superfície ou no fundo (Figura 38). O maior valor (46,57 mg.L-1) foi registrado na estação R5 e o menor (14,85 mg.L-1), na estação Z3R, ambas em Itaorna. No Saco Piraquara de Fora, as estações R3 e R4 também apresentaram valores altos na superfície (37,9 e 35,7 mg.L-1, respectivamente) e no fundo (46,20 e 43,80 mg.L-1, respectivamente). Esses valores são considerados muito elevados para a região e indicam um possível aporte de sólidos proveniente do ambiente terrestre.
Para clorofila a, as duas radiais mostraram distribuições espaciais distintas. As estações de Itaorna apresentaram valores mais elevados na superfície, enquanto que, nas estações do Saco Piraquara de Fora, os maiores valores foram registrados a 5 m de profundidade (Figura 39). O maior valor (6,62 µg.L-1) foi encontrado na estação R1 e o menor (0,20 µg.L-1), na estação R3, ambos no fundo e na mesma radial. Esse parâmetro apresentou assim uma alta variabilidade (CV 58%), sendo que os valores obtidos na campanha de 2002 foram superiores às médias registradas para a região em estudos anteriores.
Figura 38 - Distribuição espacial de MPS (mg.L-1)
Figura 39 - Distribuição espacial de clorofila a (µg.L-1)
No ciclo de 24 horas, a amostragem foi feita nos dias 28 e 29 de maio de 2002, nos pontos Z3 e Z4, com coleta de seis em seis horas (maré de enchente/alta nos períodos de 11:30-12:05h e 01:03-02:29h e maré de vazante/baixa nos períodos de 16:50-17:55h e 05:02-06:17h). Os parâmetros analisados (Tabela 36 e Tabela 37) foram os mesmos da variação espacial.
Os resultados caracterizaram as variações de temperatura como bem distintas entre os dois pontos. Na estação Z3, as temperaturas de superfície e de fundo ficaram próximas uma da outra, variando entre 23,0 e 26,0ºC, enquanto na estação Z4 os valores oscilaram entre 24,5 e 30,3ºC, mostrando estratificação (Figura 40). De uma forma geral, foi possível observar uma diminuição nos valores de temperatura a partir da primeira enchente até a segunda vazante.
Figura 40 - Variação da temperatura durante a amostragem de microvariação
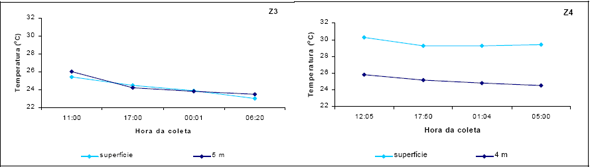
Tabela 36 - Dados coletados em maio de 2002 para avaliar a microvariação temporal e o efeito da maré, em Itaorna e no Saco Piraquara de Fora - temperatura, salinidade, oxigênio, pH, ortofosfato e fósforo total.
Estações |
Horário de coleta |
Temp.
(ºC) |
Salinidade
(S) |
Oxigênio
(mL.L-1) |
pH |
Ortofosfato
(µM P-PO4-3) |
Fósforo total
(µM P) |
sup |
*5 m |
sup |
*5 m |
Sup |
*5 m |
sup |
*5 m |
sup |
*5 m |
sup |
*5 m |
| 1 - Z3 |
11:00 |
25,4 |
26,0 |
31,9 |
32,3 |
4,34 |
4,41 |
8,08 |
8,00 |
0,23 |
0,19 |
0,75 |
1,10 |
| 2 - Z3 |
17:00 |
24,5 |
24,2 |
30,6 |
32,3 |
4,62 |
4,41 |
8,13 |
8,00 |
0,14 |
0,15 |
0,43 |
0,45 |
| 3 - Z3 |
00:01 |
23,9 |
23,8 |
31,3 |
32,8 |
1,75 |
1,68 |
8,00 |
7,90 |
0,16 |
0,20 |
0,28 |
0,70 |
| 4 - Z3 |
06:20 |
23,8 |
23,5 |
31,6 |
32,9 |
1,12 |
1,12 |
7,87 |
7,57 |
0,16 |
0,20 |
0,46 |
0,44 |
| 1 - Z4 |
12:05 |
30,3 |
25,8 |
32,3 |
32,7 |
4,76 |
4,13 |
8,00 |
8,09 |
0,20 |
0,36 |
0,58 |
0,80 |
| 2 - Z4 |
17:50 |
29,3 |
25,2 |
32,1 |
32,7 |
3,92 |
3,43 |
8,14 |
8,21 |
0,22 |
0,26 |
0,29 |
0,43 |
| 3 - Z4 |
01:04 |
29,3 |
24,8 |
32,3 |
32,7 |
1,75 |
1,61 |
8,19 |
8,00 |
0,22 |
0,21 |
0,48 |
0,51 |
| 4 - Z4 |
05:00 |
29,4 |
24,5 |
32,2 |
32,7 |
0,91 |
1,19 |
8,08 |
8,14 |
0,25 |
0,20 |
0,54 |
0,29 |
*As coletas de fundo nas estações da radial Z4 foram realizadas a 4 m de profundidade.
Tabela 37 - Dados coletados em maio de 2002 para avaliar a microvariação temporal e o efeito da maré, em Itaorna e no Saco Piraquara de Fora - nitrito, nitrato, silicato, MPS e clorofila a.
Estações |
Horário de coleta |
Amônia
(µM NH3) |
Nitrito
(µM N-NO2-) |
Nitrato
(µM N-NO3-) |
Silicato
(µM Si-SiO2) |
MPS
(mg.L-1) |
Clorofila a
(µg.L-1) |
| sup |
*5 m |
sup |
*5 m |
sup |
*5 m |
sup |
*5 m |
sup |
*5 m |
sup |
*5 m |
| 1 - Z3 |
11:00 |
1,32 |
<0,05 |
0,23 |
0,19 |
0,12 |
0,62 |
5,81 |
5,60 |
30,40 |
31,70 |
1,96 |
1,74 |
| 2 - Z3 |
17:00 |
<0,05 |
<0,05 |
0,14 |
0,15 |
<0,05 |
<0,05 |
7,07 |
3,34 |
34,50 |
32,60 |
3,71 |
2,33 |
| 3 - Z3 |
00:01 |
0,17 |
0,16 |
0,03 |
0,06 |
<0,05 |
<0,05 |
5,29 |
3,97 |
29,10 |
29,80 |
1,99 |
2,74 |
| 4 - Z3 |
06:20 |
0,06 |
0,12 |
0,05 |
0,07 |
<0,05 |
<0,05 |
4,78 |
4,90 |
29,70 |
30,20 |
1,85 |
1,99 |
| 1 - Z4 |
12:05 |
<0,05 |
0,51 |
0,02 |
0,09 |
<0,05 |
<0,05 |
4,23 |
6,04 |
25,90 |
32,60 |
0,505 |
3,47 |
| 2 - Z4 |
17:50 |
0,10 |
0,35 |
0,03 |
0,03 |
0,71 |
0,07 |
7,08 |
3,54 |
21,90 |
33,70 |
0,85 |
4,01 |
| 3 - Z4 |
01:04 |
<0,05 |
0,07 |
0,06 |
0,04 |
<0,05 |
<0,05 |
4,98 |
4,47 |
30,80 |
31,10 |
0,88 |
2,01 |
| 4 - Z4 |
05:00 |
<0,05 |
0,31 |
0,05 |
<0,02 |
<0,05 |
<0,05 |
4,12 |
3,20 |
28,20 |
30,70 |
0,22 |
2,54 |
*As coletas de fundo nas estações da radial Z4 foram realizadas a 4 m de profundidade.
Nas duas estações, os valores de salinidade foram maiores no fundo do que na superfície (Figura 41). Na estação Z3, os valores oscilaram entre 30,6 e 32,9 S e o menor valor foi responsável por uma queda brusca na salinidade de superfície, da primeira enchente para a primeira vazante; em seguida, os valores voltaram a subir. Na estação Z4 a variação foi menor (CV 1%) e os valores mais próximos entre si, oscilando entre 32,1 e 32,7 S.
Figura 41 - Variação da salinidade durante a amostragem de microvariação
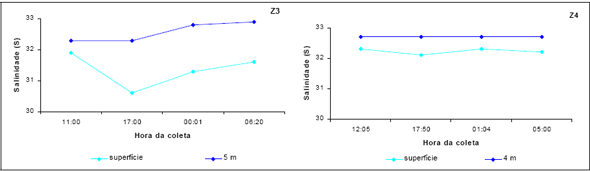
Para o oxigênio dissolvido foi observado o mesmo padrão de variação durante o ciclo de 24h. Nas duas estações, os teores de oxigênio na água sofreram uma queda brusca durante a noite, provavelmente devido à inexistência de fotossíntese e ao consumo do oxigênio só ocorrer nos processos de respiração (Figura 42). Na estação Z3, as concentrações de oxigênio dissolvido oscilaram entre 1,12 e 4,62 mL.L-1 e, na estação Z4, entre 0,91 e 4,76 mL.L-1. Tais valores são considerados normais em variações de ciclos diários, apesar de alguns deles serem inferiores ao valor mínimo estabelecido pela Resolução Conama nº 20/86 para águas salinas da classe 5 (4,20 mL.L-1), porém sempre dentro dos limites da classe 6 (2,8 mL.L-1).
Figura 42 - Variação do oxigênio dissolvido durante a amostragem de microvariação
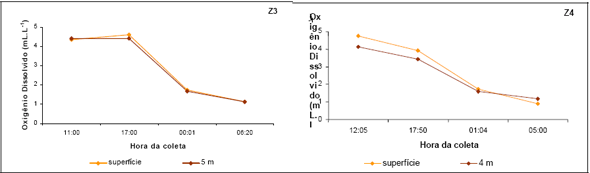
As variações do pH durante o ciclo de 24h foram bem distintas nas duas estações. Na Z3, ocorreu uma diminuição gradativa nos valores no período noturno, oscilando entre 7,57 e 8,13, e os valores no fundo foram menores que os de superfície. Na estação Z4, os valores estiveram entre 8,00 a 8,21 e, no fundo, apresentaram um aumento da primeira enchente para a primeira vazante, seguido de uma queda na segunda enchente e voltando a subir na segunda vazante; os valores de superfície apresentaram um aumento gradativo até a segunda enchente, seguido de uma leve queda na segunda vazante (Figura 43). Alguns dos valores de pH registrados foram baixos, porém dentro dos padrões aceitos para tais profundidades (Niencheski e Baumgarten 1999).
Figura 43 - Variação do pH durante a amostragem de microvariação
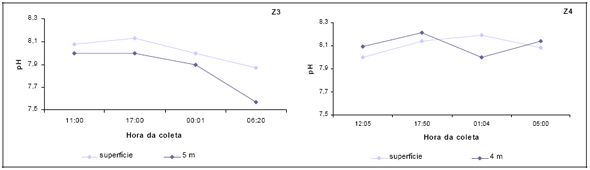
Na estação Z4, as concentrações de ortofosfato foram maiores, variando entre 0,20 e 0,36 µM, e ocorreu uma diminuição gradativa nos valores, na camada de fundo, e o inverso, na superfície. Na estação Z3, as concentrações variaram entre 0,14 e 0,23 e os valores diminuíram na primeira vazante, aumentando em seguida e permanecendo constante (Figura 44).
Figura 44 - Variação do ortofosfato durante a amostragem de microvariação
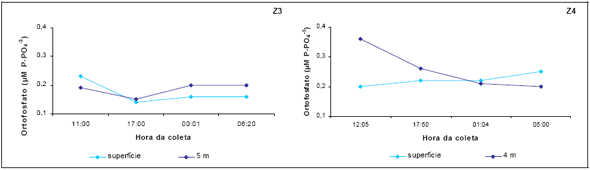
As concentrações de fósforo total variaram de 0,28 a 1,10, na estação Z3, e de 0,29 a 0,80, na estação Z4, apresentando valores maiores na camada de fundo. As duas estações apresentaram uma variação semelhante ao longo do ciclo de 24h, com exceção do valor de superfície da estação Z3, na segunda enchente, que diminuiu em relação à primeira vazante, diferentemente do observado na estação Z4 (Figura 45).
As concentrações de amônia oscilaram desde o limite de detecção do método (<0,05 µM) até 1,32 µM, na estação Z3, e até 0,51 µM, na estação Z4. Em geral, os valores foram mais elevados no fundo do que na superfície, sendo que, na estação Z4, a amônia foi detectada na superfície somente na segunda enchente, com uma concentração de 0,10 µM (Figura 46). Todos os valores observados estiveram bastante abaixo do limite estabelecido pela Resolução Conama nº 20/86 para águas da classe 5 (30 µM de N-NH3).
Figura 45 - Variação do fósforo total durante a amostragem de microvariação
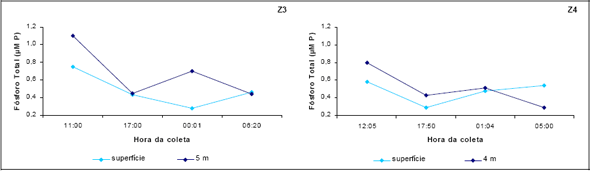
Figura 46 - Variação da amônia durante a amostragem de microvariação
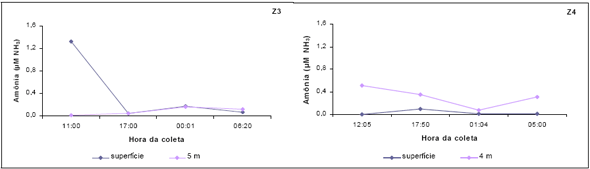
Durante o ciclo de 24h, as concentrações de nitrito diminuíram gradativamente nas duas estações, exceto na superfície da estação Z4, onde aumentaram (Figura 47). Os valores variaram de 0,03 a 0,23 µM, na estação Z4, e de não detectável (<0,02 µM) a 0,19 µM, na estação Z4.
Figura 47 - Variação do nitrito durante a amostragem de microvariação
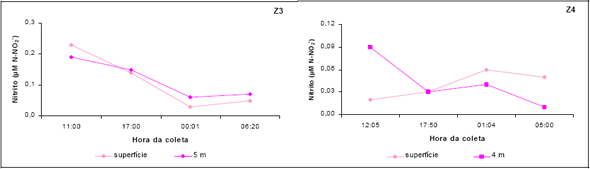
O nitrato apresentou concentrações abaixo do nível de detecção do método (0,05 µM) na maior parte das coletas (Figura 48). Em cada estação foram detectados apenas dois valores: na estação Z3, 0,12 µM na superfície, na primeira enchente, e 0,62 µM no fundo, na primeira vazante; na estação Z4, 0,71 µM na superfície, na primeira enchente, e 0,07 µM no fundo, na primeira vazante.
Figura 48 - Variação do nitrato durante a amostragem de microvariação
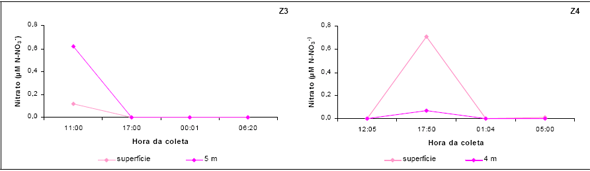
O silicato apresentou uma variação temporal semelhante nas duas estações: na superfície, ambas mostraram um aumento nas concentrações da primeira enchente para a primeira vazante, seguido de uma diminuição; na camada de fundo ocorreu o inverso, ou seja, as concentrações diminuíram da primeira enchente para a primeira vazante e em seguida aumentaram (Figura 49). As concentrações oscilaram entre 3,34 e 7,07, na estação em Z3, e entre 3,20 e 7,08, na estação Z4, e se situam dentro dos valores encontrados na variação espacial.
Figura 49 - Variação de silicato durante a amostragem de microvariação
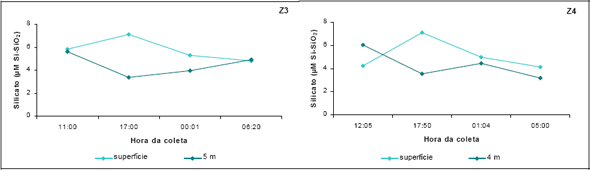
Os valores do material particulado em suspensão apresentaram uma variação temporal semelhante na camada de fundo nas duas estações, variando entre 29,8 e 32,6 na Z3 e entre 30,7 e 33,7 em Z4. Na superfície os valores foram diferentes, oscilando entre 29,1 e 34,5 na estação Z3 e entre 21,9 e 30,8 na estação Z4 (Figura 50).
Figura 50 - Variação do material particulado em suspensão durante a amostragem de microvariação
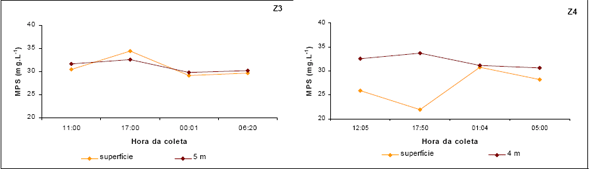
Na estação Z3, as concentrações de clorofila a no fundo e na superfície foram equivalentes, com os valores oscilando entre 1,74 e 3,71 µg.L-1; os valores na superfície foram em média maiores do que na estação Z4. Nesta última, a concentração variou entre 0,22 e 4,01 µg.L-1 e foi maior no fundo do que na superfície (Figura 51).
Figura 51 - Variação da clorofila a durante a amostragem de microvariação
7.2.1.2. Análise das Regiões de Itaorna e Piraquara de Fora - topo
As estações de Itaorna não mostraram estratificação entre a superfície e o fundo em relação à temperatura, como ocorreu nas estações do Saco Piraquara de Fora. Apesar disso, as variações nos valores obtidos para esse parâmetro nas duas estratégias de amostragem (espacial e temporal) foram muito próximas.
Quanto à salinidade, as camadas de fundo mostraram-se mais homogêneas que as de superfície, tanto na variação espacial quanto na microvariação. Além disso, como observado na análise da variação espacial, Piraquara de Fora apresentou valores mais próximos entre a superfície e o fundo do que os encontrados em Itaorna, indicando uma estratificação mais nítida nesta região.
A distribuição de oxigênio dissolvido ao longo da coluna d’água mostrou-se bem homogênea nas duas estratégias de amostragem. Na microvariação, entretanto, observou-se uma diminuição nas concentrações desse parâmetro no período noturno nas estações Z3 e Z4, devido à ausência de fotossíntese e ao consumo do oxigênio na respiração.
O pH também apresentou uma distribuição homogênea na superfície e no fundo, tanto em Itaorna quanto em Piraquara de Fora. Na microvariação, o pH diminuiu durante a noite na estação Z3 (Itaorna), mas permaneceu praticamente constante na estação Z4 (Piraquara de Fora).
Com relação aos nutrientes, as concentrações de ortofosfato foram maiores na microvariação que na variação espacial, porém as estações Z3 e Z4 apresentaram distribuição semelhante desse parâmetro. O fósforo total apresentou distribuição semelhante em Itaorna e em Piraquara de Fora, sendo que, na variação espacial, os valores foram maiores na superfície e, na microvariação, no fundo.
A amônia foi detectada na superfície somente na estação Z3, durante a microvariação. Na camada de fundo, apresentou maior concentração em Piraquara de Fora e praticamente não foi detectada em Itaorna. Ainda assim, seus valores estiveram bem abaixo do limite estabelecido pela Resolução Conama nº 20/86 para águas salinas da classe 5.
O nitrito apresentou uma diferença considerável entre as estações Z3 e Z4 na amostragem de microvariação, com a primeira apresentando valores quase duas vezes mais altos do que a segunda, tanto na superfície quanto no fundo. Na variação espacial, a distribuição nas duas regiões foi semelhante e maior na camada de fundo.
A distribuição de nitrato em Itaorna e Piraquara de Fora foi semelhante. Na variação especial, as concentrações foram, em geral, mais altas no fundo, apesar de terem sido registrados dois valores maiores na superfície. Na microvariação, esse nutriente foi detectado em apenas na primeira enchente e na primeira vazante.
O maior valor de silicato foi registrado em Itaorna e o menor, no Saco Piraquara de Fora, em ambos os casos na superfície. A concentração foi maior na superfície, nas estações de Itaorna e em algumas das estações de Piraquara de Fora. Na microvariação, a variação nas estações Z3 e Z4 foi semelhante.
O material particulado em suspensão apresentou valores altos, inclusive bem superiores aos registrados nas campanhas anteriores, indicando a ocorrência de uma condição atípica no local no momento da coleta. Não foi possível identificar qualquer padrão de distribuição desse parâmetro na coluna d’água ou entre estações. Na microvariação, na estação Z4, constatou-se uma diferença na concentração de sólidos da superfície para o fundo, porém na estação Z3 a variação foi muito semelhante nos dois estratos da coluna d’água.
Os valores obtidos para clorofila a também foram mais altos do que os registrados em campanhas anteriores, não tendo sido identificado o que motivou tal condição, aparentemente atípica. As estações de Itaorna apresentaram valores mais elevados na superfície e as de Piraquara de Fora, no fundo. Na microvariação, na superfície e no fundo, a estação Z3 apresentou variação semelhante e a estação Z4, variação distinta.
7.2.2. Fitoplâncton - topo
Os estudos disponíveis sobre o fitoplâncton da área de estudo podem ser divididos em três fases, em função do objetivo, do esforço de amostragem e da área de abrangência de cada levantamento: estudos pré-impacto, elaboração de catálogo de espécies e monitoramento.
Os estudos pré-impacto (1970-72 e 1980-81) envolveram a realização de dois levantamentos, o primeiro no período julho/70-junho/72 (Femar, 1972), tendo como objetivo analisar qualitativa e quantitativamente o fitoplâncton de Itaorna, Mamede e do Saco Piraquara de Fora. Foram feitas 12 expedições, com coletas à superfície, sendo o fitoplâncton avaliado em grandes grupos taxonômicos (divisão, classe e família), sem detalhamento em nível específico. O segundo levantamento foi realizado no período janeiro/80-janeiro/81 e compreendeu campanhas mensais, com coletas realizadas na superfície, a 5 m de profundidade e na profundidade de Secchi, tendo como objetivo analisar qualitativa e quantitativamente o fitoplâncton de Itaorna (captação de água), do Saco Piraquara de Fora (lançamento do efluente) e do Saco Piraquara de Dentro (área de controle). Esse levantamento foi promovido em convênio por Furnas e a Fundação José Bonifácio e executado pelo Instituto de Biologia da Universidade Federal do Rio de Janeiro (FUJB, 1981), tendo gerado três trabalhos sobre a abundância e a composição das populações fitoplanctônicas: Bradford (1983), Villac (1983) e Petretski (1987).
A elaboração do catálogo de espécies (março/89-agosto/91) envolveu a realização de campanhas no período outubro-dezembro/89, nos meses de março, abril, junho, julho, agosto e outubro/90 e nos meses de março, abril, maio, julho e agosto/91 e compreendeu arrastos verticais (de 5 m de profundidade à superfície) com rede de 50µm de malha, nos pontos Z3, Z4 e 47B. Os organismos foram estudados (morfometria), identificados e fotografados para a confecção do catálogo (Tenenbaum et al., 1991).
O monitoramento foi iniciado em outubro/87, a partir da implantação do Central Nuclear Almirante Álvaro Alberto (CNAAA), e seus resultados constam de 13 relatórios técnicos anuais contendo os dados obtidos até dezembro/01 (Tenenbaum & Villac, 1990, Tenenbaum et al., 1991, Tenenbaum et al., 1992, Tenenbaum & Nascimento, 1993, Tenenbaum et al., 1994, Tenenbaum & Dias, 1995, Tenenbaum & Villac, 1996, Tenenbaum et al., 1997, Tenenbaum et al., 1998, Tenenbaum & Villac, 1999, Tenenbaum et al., 2000, Tenenbaum et al., 2001, Tenenbaum et al., 2002). Desde o início do monitoramento vêm sendo empregadas as mesmas metodologias de amostragem e de análise.
Quanto aos aspectos qualitativos, esses estudos registraram 373 táxons, distribuídos em diatomáceas (238), dinoflagelados (118), cocolitoforídeos (5), silicoflagelados (3), cianofíceas (3), euglenofíceas (2), ebriideas (2), clorofíceas (1) e prasinofíceas (1). Constatou-se a predominância de espécies do microfitoplâncton (organismos superiores a 20 µm) influenciada, principalmente, pela metodologia empregada.
Os organismos nanoplanctônicos, principalmente os flagelados, precisam ser avaliados por metodologia diferente, em função de sua faixa dimensional, e preferencialmente vivos, devido a alterações morfológicas provocadas pela fixação da amostra.
A composição da comunidade fitoplanctônica da área de estudo apresenta, quantitativa e qualitativamente, contribuição marcante de diatomáceas (Tabela 38), uma característica de regiões costeiras. Algumas espécies indicam a presença de água de plataforma, várias delas com distribuição cosmopolita e típicas de regiões tropicais e subtropicais.
Tabela 38 - Fitoplâncton - Estudos Anteriores - Número de espécies por classe taxonômica
| Discriminação |
Constante
>_ 75% dos estudos |
Freqüente
50-75% dos estudos |
Presente
25-50% dos estudos |
Rara
< 25% dos estudos |
| Diatomáceas |
36 |
21 |
41 |
123 |
| Dinoflagelados |
6 |
17 |
17 |
75 |
| Cocolitoforídeos |
0 |
0 |
2 |
3 |
| Silicoflagelados |
1 |
0 |
0 |
2 |
| Cianofíceas |
0 |
0 |
1 |
2 |
| Euglenofíceas |
0 |
0 |
0 |
2 |
| Ebriideas |
1 |
0 |
0 |
1 |
| Clorofíceas |
0 |
0 |
0 |
1 |
| Prasinofíceas |
0 |
0 |
0 |
1 |
Em termos quantitativos, a densidade celular do fitoplâncton da região vem oscilando entre 103 e 106 células por litro, com valores médios em torno de 104-105 células por litro (Tabela 39), sendo importante destacar que a comparação entre os dados da fase de pré-funcionamento das usinas com os da fase de monitoramento deve ser feita com cautela. Foram considerados apenas os estudos nos quais a coleta foi feita com garrafa e análise qualitativa e quantitativa, pelo método de sedimentação (n= número de amostras, total=nano + microfitoplâncton, micro=>20µm).
Tabela 39 - Fitoplâncton - Estudos Anteriores - Densidade celular (células/L)
Pré-funcionamento (1980) |
Monitoramento (1987-2001) |
Densidade |
Itaorna
n=33 |
Piraquara de Fora
n=68 |
Z3
n= 162 |
Z4
n= 161 |
47B
n= 162 |
| Total |
Micro |
Total |
Micro |
Total |
Micro |
| Mínima |
1.240 |
960 |
19.372 |
128 |
10.400 |
700 |
14.900 |
840 |
| Máxima |
238.702 |
321.875 |
5.069.460 |
4.670.360 |
3.593.650 |
3.428.550 |
2.327.498 |
2.154.598 |
| Média |
15.862 |
22.743 |
344.447 |
130.862 |
259.941 |
89.598 |
273.385 |
96.642 |
| Desvio |
40.603 |
41.634 |
56.798 |
463.709 |
397.237 |
315.696 |
380.519 |
272.272 |
A metodologia empregada em 1980 considera principalmente os organismos da fração do microfitoplâncton (organismos > 20µm), mas inclui algumas espécies importantes (Skeletonema costatum) consideradas, atualmente, na fração do nanoplâncton (organismos £ 20µm). Essa ordem de grandeza é considerada nos estudos como característica de região costeira sem uma fonte importante de eutrofização natural (rios de grande porte ou ressurgência).
A região vem sendo considerada espacialmente homogênea desde o período de pré-funcionamento das usinas, sem diferenças marcantes na composição e na abundância do fitoplâncton entre Itaorna e Piraquara de Fora (Petretsky, 1987; Tenenbaum & Villac, 1996). Observa-se, entretanto, uma variação sazonal nítida, tanto em termos de riqueza específica (número de espécies) quanto em relação à abundância dos organismos. Esse padrão não é claro no período pré-funcionamento das usinas, que conta com apenas um ano de dados e inclui organismos do nanoplâncton na análise, o que torna a tendência menos marcante. Os dados referentes ao período de monitoramento (1987-2001) monstram essa sazonalidade de maneira mais nítida (Figura 52).
As variações sazonais são regidas, principalmente, pela climatologia da região, estabelecendo uma seqüência de sucessão. Postula-se que o verão quente e chuvoso representa o impacto ambiental (natural) a partir do qual a sucessão se inicia e evolui com a maior estabilidade ambiental do resto do ano. É no verão que se observam os valores de clorofila e densidade celular mais altos. O meio-final do outono representa um período de menor produtividade fitoplanctônica (densidades mais baixas e menor número de espécies), quando o ambiente parece estar exaurido após um verão de intensa atividade. Durante um inverno ameno, as populações fitoplanctônicas começam a se desenvolver, inclusive com uma maior disponibilidade de silicato, elemento indispensável para a reprodução das diatomáceas, como demonstrado na Figura 52e.
Figura 52 - Fitoplâncton - Estudos Anteriores - Variação espacial e temporal da riqueza específica e da abundância (OBS.: n entre 10 e 15)
O número de espécies é bem maior na primavera, enquanto que a maior abundância é observada no período primavera-verão. A comparação entre três indicadores de abundância (clorofila a, densidade celular total e densidade celular do microfitoplâncton) utilizados no período de monitoramento evidencia que os organismos de maior porte são os sensores mais sensíveis das variações ambientais da área de estudo (a clorofila a inclui organismos >0,45µm, o fitoplâncton total os >2µm e o microfitoplâncton os >20µm).
A primavera apresenta um aumento na densidade celular (e consumo de silicato), mas com uma estrutura de comunidade mais madura, com diatomáceas de grande porte e, em alguns anos, dinoflagelados do microfitoplâncton. O verão apresenta alta densidade celular causada por espécies de pequeno porte, mas que pode estar associada a valores mais altos de riqueza específica devido a um efeito do incremento de espécies remanescentes da primavera.
No entanto, apesar da grande riqueza de espécies observada freqüentemente na primavera, a interrupção na seqüência da sucessão pode ocorrer devido à dominância de diatomáceas de pequeno porte, como Cylindrotheca closterium, Dactyliosolen fragilissimus, Leptocylindrus minimus, Pseudo-nitzschia delicatissima. Essas espécies podem ser consideradas oportunistas, associadas aos primeiros estágios de sucessão fitoplanctônica (sensu Margalef, 1958), que respondem rapidamente a algum tipo de interferência ambiental.
Durante os 16 anos de monitoramento, as sínteses anuais da dinâmica fitoplanctônica, elaboradas a partir de análise multivariada (ordenação e agrupamento), identificam os fatores climáticos (pluviosidade e temperatura) como os mais importantes na formação das tendências observadas, pelo fato de desencadearem o mecanismo de sucessão antes descrito. Entretanto, observam-se alterações nesses padrões quando o mecanismo de sucessão é bruscamente interrompido e/ou reiniciado, em um momento inesperado do ano, geralmente por diatomáceas oportunistas.
As evidências apresentadas nos estudos permitem especular acerca dos mecanismos de caráter natural e/ou antrópico responsáveis por tais alterações. A ecologia dos sistemas marinhos está sujeita a variações associadas ao fenômeno El Niño, com conseqüências notáveis para o fitoplâncton (Barber & Chavez, 1983). No período dos estudos, o fenômeno ocorreu em duas ocasiões, de 1991 a 1995 e em 1998, tendo alterado em ambos os casos os padrões climáticos.
O mecanismo antrópico, associado ao funcionamento da CNAAA, estaria relacionado ao aumento da temperatura da água e ao turbilhonamento em Piraquara de Fora, assim como a dragagens realizadas em Itaorna, com efeitos mais agudos após a ativação da unidade 2. De fato, desde 2000, vem sendo observada uma diminuição marcante na densidade celular e na riqueza específica, associada à presença de detritos inorgânicos nas amostras e à diminuição da transparência da água. A Figura 54 apresenta a série histórica (1995-2000) dos valores mínimos, máximos e médios de densidade celular total dos três pontos de coleta do monitoramento, verificando-se que, em 2000, a densidade celular média foi de uma ordem de grandeza inferior à dos anos anteriores, coincidindo com os valores mínimos observados.
Figura 53 - Influência do El Niño na pluviosidade e na temperatura
Figura 54 - Fitoplâncton -Estudos Anteriores - Densidade celular no período 1995-2000
Para permitir a comparação com os dados obtidos nos estudos anteriores, foram empregadas, na campanha de 2002, as mesmas metodologias de amostragem e de análise adotadas desde o início do monitoramento (outubro/87), tendo sido analisada apenas a fração do microfitoplâncton (organismos > 20µm), tendo em vista que, conforme evidenciado pelos dados disponíveis, esses organismos se mostraram mais sensíveis às variações ambientais detectadas.
Em todos os pontos foram coletadas amostras de água na superfície e a 5 m de profundidade, por amostragem radial e por microvariação segundo diferentes horários de maré, totalizando 40 amostras (Tabela 40). A coleta foi feita com garrafa de Van Dorn, da qual foram retirados e acondicionados em frascos de polietileno 250 mL para análise qualitativa e quantitativa do fitoplâncton, sendo os organismos imediatamente fixados em formaldeído tamponado com tetraborato de sódio a uma concentração final de 2% (modificado de Sournia, 1978).
Tabela 40 - Fitoplâncton - Campanha de 2002 - Identificação das amostras coletadas
Método |
Ponto de coleta |
Data |
Hora |
Radial |
R1 |
27/05/2002 |
15:23-16:00 |
| R2 |
27/05/2002 |
16:05-16:32 |
| R3 |
27/05/2002 |
17:03-17:20 |
| R4 |
27/05/2002 |
17:31 |
| R5 |
27/05/2002 |
11:40-12:10 |
| R6 |
27/05/2002 |
12:15-12:50 |
| R7 |
27/05/2002 |
12:57-13:15 |
| R8 |
27/05/2002 |
13:25-13:40 |
| R9 |
27/05/2002 |
13:55-14:15 |
| Z3R |
27/05/2002 |
10:35-11:30 |
| Z4R |
27/05/2002 |
14:56-15:15 |
| 47BR |
27/05/2002 |
16:37-16:55 |
Microvariação (*) |
1Z3 |
28/05/2002 |
11:00-11:30 |
| 2Z3 |
28/05/2002 |
16:50-17:15 |
| 3Z3 |
29/05/2002 |
00:02-00:29 |
| 4Z3 |
29/05/2002 |
06:17-06:46 |
| 1Z4 |
28/05/2002 |
12:05-12:46 |
| 2Z4 |
28/05/2002 |
17:55-18:37 |
| 3Z4 |
29/05/2002 |
01:03-01:40 |
| 4Z4 |
29/05/2002 |
05:02-05-35 |
(*) Os números 1 e 3 que antecedem o ponto de coleta indicam coletas em maré de enchente/alta e os números 2 e 4, coletas em maré vazante/baixa (maré de sizígia, com variação ente 0,1 e 1,2 m.
Em laboratório, as análises qualitativa e quantitativa foram feitas pelo método de sedimentação (Utermöhl, 1958), sendo os organismos da fração do microfitoplâncton (células > 20µm) quantificados e identificados, quando possível, em nível de espécie. A análise foi realizada em meia câmara de contagem, com aumento de 200X (duzentas vezes), utilizando cubetas de 50 cm3, com período de sedimentação entre 40 e 48 horas. A ocorrência das espécies foi calculada considerando as 40 amostras e classificadas em categorias arbitrárias, conforme apresentado na Tabela 41.
Tabela 41 - Categorias de classificação de ocorrência de espécies de fitoplâncton.
Categoria |
Ocorrência |
| Rara |
Em £ 10 % das amostras |
| Presente |
Em >10 % e £ 50 % das amostras |
| Freqüente |
Em > 50% e £ 80 % das amostras |
| Constante |
Em > 80 % das amostras |
As espécies dominantes e/ou abundantes de cada amostra foram determinadas segundo a concepção de Lobo & Leighton (1986), entendendo-se como espécie dominante aquela cuja densidade supera 50% da densidade total e, como espécie abundante, aquela cuja densidade supera a densidade média dos organismos considerados no cálculo. Para a amostragem radial foram elaborados mapas de distribuição (isolinhas) com o emprego do programaSurfer.
7.2.2.1. Caracterização da Comunidade Fitoplanctônica- topo
O inventário de espécies registrou 96 táxons (Tabela 42), distribuídos em diatomáceas (55), dinoflagelados (35), silicoflagelados (2), ebriideas (2), cianofíceas (1) e cocolitoforídeos (1). Foram encontradas também euglenofíceas, não identificadas em nível específico.
Tabela 42 - Fitoplâncton - Campanha de 2002 - Lista de espécies
Essa composição florística, com predomínio de diatomáceas, é compatível com a observada nos estudos anteriores, sendo característica de regiões costeiras (Barnes & Hughes, 1982). O número de espécies encontradas no levantamento das radiais (10) foi ligeiramente superior ao observado no estudo de microvariação, o que se justifica pela diferença de esforço amostral, com maior representatividade espacial do primeiro. As novas ocorrências observadas foram a diatomácea Coscinodiscus cf. granii e os dinoflagelados Ceratium furca var. eugrammum, Cladopyxis sp. (gênero ainda não encontrado na região), Paleophalacroma verrucosum e cf. Warnowia fusus.
A densidade celular do microfitoplâncton total de todo o período de coleta (27-29 de maio de 2002) variou entre 9,9 x 103 e 8,3 x104 cel.L-1 (Tabela 43). Essa ordem de grandeza está de acordo com o observado nos estudos anteriores (Figura 52), mais especificamente com o período outono-inverno, quando foram observadas as concentrações fitoplanctônicas mais baixas do ano. Como esperado, as diatomáceas constituíram a classe taxonômica mais abundante (média: 2,6 x 104 ± 1,4 x 104 cel.L-1), seguida pelos dinoflagelados (média: 1,9 x 103 ± 4,7 x 103 cel.L-1).
Tabela 43 - Microfitoplâncton - Campanha de 2002 - Estatística descritiva da variação da densidade celular do conjunto de dados
Discriminação |
Conjunto de dados (n=40) |
| Mínimo |
Máximo |
Média |
Desvio |
| cél.L-1 |
% |
cél.L-1 |
% |
cél.L-1 |
% |
cél.L-1 |
% |
| Diatomáceas |
9.240 |
61,53 |
78.567 |
99,55 |
26.823 |
94,27 |
14.122 |
8,79 |
| Dinoflagelados |
80 |
0,42 |
28.092 |
38,41 |
1.939 |
5,32 |
4.718 |
8,84 |
| Cianofíceas |
nd |
- |
200 |
0,86 |
44 |
0,17 |
52 |
0,21 |
| Silicoflagelados |
nd |
- |
120 |
0,57 |
11 |
0,05 |
30 |
0,14 |
| Ebriideae |
nd |
- |
160 |
1,20 |
29 |
0,12 |
42 |
0,23 |
| Euglenofíceas |
nd |
- |
520 |
0,62 |
24 |
0,05 |
84 |
0,13 |
| Cocolitoforídeos |
nd |
- |
80 |
0,35 |
3 |
0,01 |
14 |
0,06 |
| Total |
9.960 |
|
83.487 |
|
28.875 |
|
16.124 |
|
nd = não detectável.
Algumas espécies se destacaram em termos de ocorrência e abundância (Tabela 44), especialmente as diatomáceas, em sua maioria células de forma alongada que, apesar de serem organismos de grande porte, alguns atingindo comprimento superior a 100µm, apresentam alta relação superfície:volume, o que lhes confere alta capacidade de absorção de nutrientes e adaptabilidade a ambientes com disponibilidade de nutrientes (meso a eutrófico) e com turbulência (Margalef, 1978; Reynolds, 1988). Deve ser dada atenção especial às espécies do gênero Pseudo-nitzschia (constante e dominante), visto que das 23 espécies descritas até o presente nove são potencialmente tóxicas (Bates, 1998; Fryxel & Villac, 1999; Sarno, 2000).
Entre os dinoflagelados encontrados na campanha de 2002, destacam-se espécies registradas anteriormente como formadoras de florações, como P. micans (Fukuyo et al., 1990), Scrippsiella sp. (Odebrecht et al., 2002) e Ceratium hircus (Steidinger & Williams, 1970).
A espécie E. tripartita é comum na região de estudo, especialmente em épocas de alta disponibilidade de sílica, pois seu exoesqueleto é essencialmente constituído por este nutriente. A cianofícea J. pellucida também é comum na área de estudo, sendo encontrada em vários ambientes da costa brasileira, de salobro a marinho, como lagoas costeiras e regiões estuarinas dos estados de Pernambuco, do Rio de Janeiro, de São Paulo e do Rio Grande do Sul (Azevedo et al., 1999).
Os resultados obtidos no levantamento das radiais e no estudo de microvariação estão sintetizados na Tabela 45 e detalhados nos itens subsequentes.
Tabela 44 - Fitoplâncton - Táxons que se destacam pela ocorrência e a contribuição quantitativa
| Táxon |
Ocorrência (a) |
Densidade (b) |
| Diatomáceas |
|
|
| Chaetoceros peruvianus |
Presente |
|
| Corethron criophilum |
Freqüente |
|
| Cylindrotheca closterium |
Constante |
Abundante |
| Fragilariineae |
Constante |
|
| Haslea wawrikae |
Constante |
|
| Pseudo-nitzschia “delicatissima” |
Constante |
Dominante |
| Pseudo-nitzschia “seriata” |
Presente |
|
| Rhizosolenia pungens |
Freqüente |
|
| Thalassionema cf. nitzschioides |
Freqüente |
|
| Thalassiothrix gibberula |
Presente |
|
| Dinoflagelados |
|
|
| Ceratium falcatum |
Presente |
|
| Ceratium hircus |
Presente |
Abundante |
| Gymnodiniaceae |
Presente |
|
| Prorocentrum gracile |
Freqüente |
|
| Prorocentrum micans |
Presente |
|
| Scrippsiella sp. |
Rara |
Abundante |
| Ebriideae |
|
|
| Ebria tripartita |
Presente |
|
| Cianofícea |
|
|
| Johannesbaptistia pellucida |
Presente |
|
(a) Rara: em <10% das amostras; presente: >10% e £50%; freqüente: >50% e £80%; constante: >80%.
(b) Dominante: densidade > 50% da amostra; abundante: densidade > densidade média da amostra.
Tabela 45 - Microfitoplâncton- Campanha de 2002 - Densidade celular na amostragem das radiais e na microvariação.
Discriminação |
Mínimo |
Máximo |
Média |
Desvio |
| cél.L-1 |
% |
cél.L-1 |
% |
cél.L-1 |
% |
cél.L-1 |
% |
Radiais (n=24) |
| Diatomáceas |
9.240 |
79,63 |
78.567 |
99,42 |
29.193 |
96,11 |
16.485 |
4,27 |
| Dinoflagelados |
80 |
0,42 |
8.840 |
20,28 |
1.205 |
3,48 |
1.891 |
4,25 |
| Cianofíceas |
nd |
- |
200 |
0,62 |
40 |
0,14 |
53 |
0,17 |
| Silicoflagelados |
nd |
- |
120 |
0,46 |
7 |
0,03 |
25 |
0,10 |
| Ebriideas |
nd |
- |
160 |
1,20 |
42 |
0,18 |
49 |
0,28 |
| Euglenofíceas |
nd |
- |
520 |
0,62 |
30 |
0,05 |
106 |
0,14 |
| Cocolitoforídeos |
nd |
- |
80 |
0,35 |
3 |
0,01 |
16 |
0,07 |
| Total |
9.960 |
|
83.487 |
|
30.521 |
|
17.465 |
|
Microvariação (n=16) |
| Diatomáceas |
9.320 |
61,53 |
44.997 |
99,55 |
23.270 |
91,50 |
8.906 |
12,62 |
| Dinoflagelados |
120 |
0,45 |
28.092 |
38,41 |
3.041 |
8,08 |
7.087 |
12,72 |
| Cianofíceas |
nd |
- |
160 |
0,86 |
50 |
0,22 |
52 |
0,26 |
| Silicoflagelados |
nd |
- |
120 |
0,57 |
18 |
0,09 |
36 |
0,18 |
| Ebriideas |
nd |
- |
40 |
0,19 |
10 |
0,04 |
18 |
0,07 |
| Euglenofíceas |
nd |
- |
120 |
0,40 |
15 |
0,06 |
32 |
0,12 |
| Cocolitoforídeos |
nd |
- |
40 |
0,13 |
3 |
0,01 |
10 |
0,03 |
| Total |
10.520 |
|
73.129 |
|
26.406 |
|
14.052 |
|
nd= não detectável.
A distribuição espacial dos estudos das radiais do microfitoplâncton (Figura 55a) mostrou valores de densidade celular em Itaorna superiores aos encontrados em Piraquara de Fora, especialmente na superfície. Em Itaorna, nas duas profundidades analisadas, foi detectado um núcleo de maior concentração fitoplanctônica nas estações R5 e R6, a partir do qual foram observadas densidades menores, em direção tanto do continente quanto do mar aberto. Em Piraquara de Fora, a superfície mostrou-se homogênea e com os menores valores observados durante a campanha; a 5 m de profundidade foi detectado um gradiente de concentrações decrescentes do continente (estação Z4) para fora da área de Piraquara (estação R4).
A riqueza de espécies por amostra (Figura 55b) pode ser considerada baixa para a área de estudo, padrão esperado para o outono (Figura 52). Em Itaorna, as camadas de superfície e a 5 m de profundidade apresentaram tendência análoga: maior riqueza mais próximo à costa (pequeno incremento de diatomáceas) e no ponto de coleta mais afastado (pequeno incremento de dinoflagelados). Em Piraquara de Fora foi detectado pequeno aumento no número de diatomáceas em R4, o ponto mais afastado do local de lançamento do efluente da usina: a 5 m de profundidade, o gradiente observado deve ser desconsiderado, por representar uma variação muito pequena (entre 7 e 11 espécies).
A distribuição espacial da densidade celular antes descrita foi determinada pela distribuição das diatomáceas (Figura 56a), mais especificamente do complexo Pseudo-nitzschia “delicatissima”, organismo considerado dominante (>50% da densidade celular da amostra) em todas as amostras analisadas. Investir no detalhamento da composição específica deste gênero com a utilização de técnicas microscópicas específica (microscopia eletrônica) é fundamental por se tratar de espécies potencialmente tóxicas, como discutido anteriormente.
A contribuição quantitativa dos dinoflagelados (Figura 56b) apresentou tendência diferente entre as áreas: crescente em Itaorna, a partir do continente (principalmente na superfície), e decrescente em Piraquara, a 5 m de profundidade. Tanto para diatomáceas quanto para dinoflagelados, Piraquara de Fora se mostrou homogênea em superfície.
Cada espécie de microalga pode apresentar limites definidos de afinidades e tolerâncias quanto a variações ambientais. Há inúmeros estudos experimentais que buscam compreender essa dinâmica, porém eles são insuficientes frente ao número de espécies de fitoplâncton existentes, estimado em 498 ± 15 gêneros e 3.910 ± 465 espécies (SOURNIA et al, 1991). A compreensão das preferências e tolerâncias das populações pode ser avaliada também a partir do papel ecológico de grupos de organismos que apresentam requisitos ambientais análogos, como o fato de as diatomáceas serem mais costeiras (essencialmente fotoautotróficas) e os dinoflagelados predominarem em águas mais oligotróficas (presença de heterotrofia facultativa) (Lalli & Parsons, 1993).
Figura 55 - Microfitoplâncton - Distribuição espacial da densidade celular (a) e da riqueza de espécies (b) entre as áreas de estudo. Obs.: profundidades de coleta com escalas diferentes para os valores de densidade celular e riqueza de espécies.
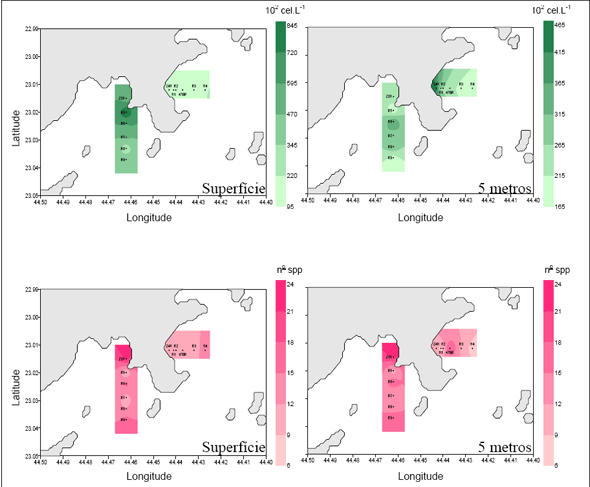
Figura 56 - Ditomáceas (a) e Dinoflagelados (b) - Distribuição espacial da densidade celular entre as áreas de estudo. Obs.: profundidades de coleta com escalas de cor diferentes para os valores de densidade celular.
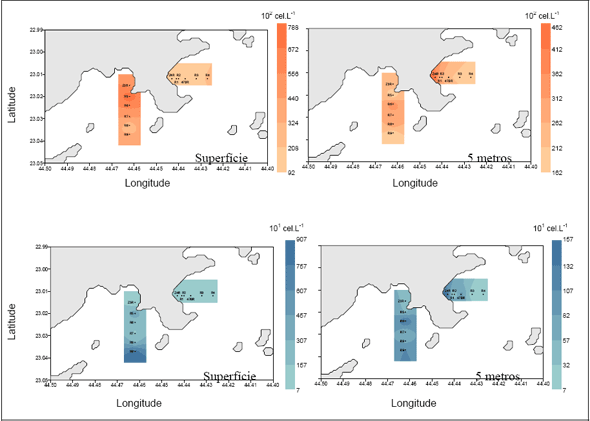
A abordagem de grupos morfofuncionais (Margalef, 1978; Reynolds, 1988) leva em consideração que a dinâmica fitoplanctônica pode ser compreendida através do estudo da relação superfície:volume das células, associada à capacidade de absorção de nutrientes, e de adaptações para manutenção da população na camada fótica (presença de flagelos, formação de cadeias de células). Desta forma, a dinâmica do fitoplâncton no ambiente pode ser estudada segundo gradientes ambientais que congregam condições específicas de disponibilidade de nutrientes e turbulência/estabilidade da coluna d’água. Esta abordagem foi aplicada, em caráter preliminar, aos dados das radiais, demonstrando uma estratificação horizontal nítida, tanto em Itaorna quanto em Piraquara de Fora (Figura 57).
Figura 57 - Distribuição espacial da relação entre diatomáceas longas e flagelados entre as áreas de estudo (Profundidades de coleta com escalas diferentes)
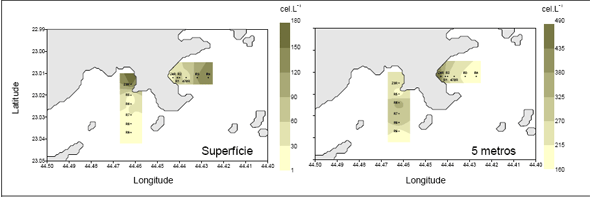
Microvariação Temporal (Maré)
As diatomáceas alongadas são mais adaptadas a ambientes com disponibilidade de nutrientes (meso a eutrófico) e maior turbulência e os flagelados (em sua maioria, dinoflagelados) possuem a vantagem de se adaptarem a ambientes com menos nutrientes e verticalmente mais estáveis. É interessante observar que o gradiente horizontal de Piraquara de Fora à superfície foi o inverso da verificada a 5 m de profundidade. Estudos futuros devem considerar a variação de turbulência em Piraquara, tanto a causada pela presença da pluma térmica quanto a definida pela variação da profundidade local (Z4 mais rasa e, portanto, com maior turbulência a 5 m de profundidade).
O fitoplâncton apresentou variações quantitativas e qualitativas no período de 24 horas (Figura 58), mais acentuadamente em Piraquara de Fora e a 5 m de profundidade. Não foi possível associar a variação à oscilação da maré, porém as maiores densidades (totais e de diatomáceas, o grupo dominante) foram observadas no final do dia, indicando que a luz é um fator controlador do fitoplâncton local.
A maior densidade celular no final do dia pode ser interpretada como o resultado da produção fitoplanctônica no período de maior insolação. De fato, no processo de divisão celular, especialmente de diatomáceas, observa-se primeiramente o aumento de pigmentos pela intensificação da atividade fotossintética, seguido do aumento de biovolume que antecede a divisão celular e, por fim, o reflexo do aumento populacional em termos de densidade celular (Jimenez et al., 1987).
Durante o período das 24 horas de coleta foi observado o surgimento, a abundância e o quase desaparecimento do dinoflagelado Ceratium hircus, especialmente em Piraquara de Fora a 5 m de profundidade (pico de abundância dos dinoflagelados na Figura 58). Como mencionado anteriormente, trata-se de espécie responsável por florações (Steidinger & Williams, 1970). Seria prematuro interpretar essa seqüência de eventos, pois a coleta foi interrompida antes de se verificar se o leve aumento observado em Itaorna resultaria em outro pico de abundância.
O estudo de microvariação foi importante para demonstrar a variabilidade temporal das populações fitoplanctônicas em curta escala de tempo. A interpretação de dados de monitoramento deve levar em conta essa questão. Somente uma série temporal de longa escala (anos) poderá minimizar tais diferenças e extrair tendências gerais para o fitoplâncton local.
Figura 58 - Variação da densidade celular do microfitoplâncton, de diatomáceas e de dinoflagelados durante a amostragem de microvariação. Maré enchente/alta nos períodos de 11:30-12:05 e 01:03-02:29. Maré vazante/baixa nos períodos de 16:50-17:55 e 05:02-06:17.
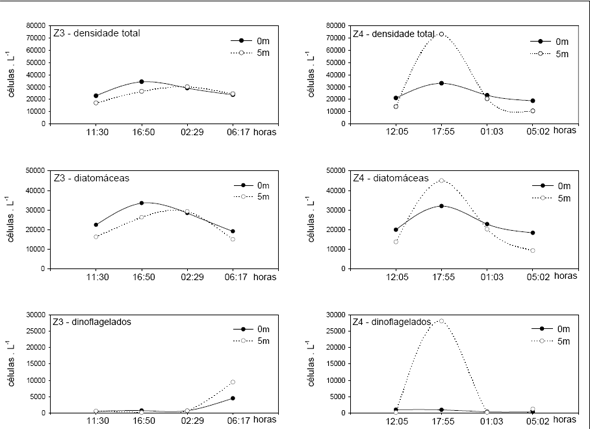
7.2.2.2. Análise das Regiões de Itaorna e Piraquara de Fora - topo
Os dados disponíveis indicaram que a região de estudo é homogênea em termos hidrológicos e da dinâmica fitoplanctônica, sem diferenças marcantes entre os pontos de coleta de Itaorna (Z3) e de Piraquara de Fora (Z4 e 47BR) (p.e., Petretsky, 1987), e que, após a ativação de Angra 2, houve uma diminuição na densidade celular, especialmente da fração do microfitoplâncton (p.e., Tenenbaum et al., 2001).
Os resultados da campanha de 2002 mostraram diferenças na distribuição espacial das populações fitoplanctônicas, seja de uma estação de coleta para outra, seja considerando cada área isoladamente. Em Itaorna, as populações de superfície e de 5 m de profundidade apresentaram um comportamento análogo, enquanto que, em Piraquara de Fora, foi detectada estratificação vertical entre o fitoplâncton de superfície e o de 5 m de profundidade. Em termos de distribuição horizontal, as diferenças observadas em Itaorna eram esperadas, pois a malha amostral utilizada adicionou pontos muito distantes do ponto monitorado (Z3), afastados da linha de costa. Já em Piraquara de Fora, a homogeneidade em superfície e o gradiente a 5 m de profundidade são resultados que alteram (e complementam) a tendência conhecida para essa enseada.
Em termos de composição florística, as duas regiões apresentaram o mesmo núcleo de espécies principais, variando apenas quanto a algumas espécies consideradas como raras na campanha de 2002 (Tabela 46). Em Itaorna (riqueza total: 70 espécies), a influência mais efetiva de águas da plataforma foi indicada pelo incremento de algumas espécies de dinoflagelados, enquanto que as águas mais rasas de Piraquara de Fora (riqueza total: 69 espécies) permitiram a ressuspensão de espécies de diatomáceas associadas ao sedimento.
Tabela 46 - Micorfictoplânton - Estatística descritiva da variação da densidade celular e da riqueza de espécies de todo o conjunto de dados.
Local |
Parâmetro |
Uni-dade |
Superfície (n=10) |
5 m de profundidade (n=10) |
| Mín |
Máx |
Média |
Desvio |
Mín |
Máx |
Média |
Desvio |
Itaorna |
Densidade celular |
cél.L-1 |
22.880 |
83.487 |
42.543 |
19.355 |
16.760 |
37.120 |
25.660 |
6.642 |
| Riqueza de espécies |
N. Spp |
12 |
21 |
16 |
3 |
7 |
23 |
15 |
5 |
Piraraquara de Fora |
Densidade celular |
cél.L-1 |
9.960 |
33.080 |
19.532 |
6.683 |
10.520 |
73.129 |
27.764 |
18.597 |
| Riqueza de espécies |
N. Spp |
8 |
25 |
14 |
5 |
5 |
19 |
11 |
4 |
Se a diferença na composição florística entre Itaorna e Piraquara de Fora é pequena, a variação em termos quantitativos é marcante (Tabela 46). A grande diferença de densidade celular de superfície observada entre Itaorna (média: 4,2 x 104 ± 1,9 x 104 cel.L-1) e Piraquara de Fora (média: 1,9 x 104 ± 6,6 x 104 cel.L-1) deve ser monitorada.
Efeitos negativos de efluentes térmicos em comunidades fitoplanctônicas podem incluir diminuição de clorofila a (Kozasa & Anraku, 1981), de densidade celular (Briand, 1975) e da taxa fotossintética (Domotor et al., 1982). A possibilidade de aclimatação de organismos fitoplanctônicos ao aumento de temperatura já foi observada em termos de modificações da taxa fotossintética (Lei & Morris in Cravens et al., 1983). Entretanto, alterações na composição específica da comunidade podem ocorrer por seleção, a favor de espécies termófilas, modificando padrões sazonais de sucessão de espécies (Carpenter, 1973; Bourgade, 1977; Simm, 1988b). Em regiões de baixa latitude, os efeitos do aquecimento podem ser mais acentuados, devido à presença de um maior número de espécies estenotérmicas que já se encontram próximas do limite máximo de temperatura (Barnett, 1972).
A homogeneidade em superfície e o gradiente a 5 m de profundidade observados em Piraquara de Fora indicam que a malha amostral deve ser expandida, incluindo duas profundidades e pontos além do R4, de modo a melhor captar a extensão da pluma térmica.
Na campanha de 2002, o inventário de espécies registrou 96 táxons: 55 diatomáceas, 35 dinoflagelados, dois silicoflagelados, dois ebriideas, uma cianofícea e um cocolitoforídeo, incluindo cinco ocorrências não registradas nas campanhas anteriores: a diatomácea Coscinodiscus cf. granii e os dinoflagelados Ceratium furca var. eugrammum, Cladopyxis sp. (gênero ainda não registrado para a região), Paleophalacroma verrucosum e cf. Warnowia fusus.
A densidade celular do microfitoplâncton do período de estudo variou de 9,9 x 103 a 8,3 x 104 cel.L-1 e a riqueza específica, de cinco a 25 espécies por amostra. Esses resultados estão em consonância com o observado anteriormente na área de estudo no outono, período do ano no qual as concentrações e a riqueza fitoplanctônicas se apresentam mais baixas. As diatomáceas constituíram a classe mais abundante (média: 2,6 x 10-1± 1,4 x 104 cel.L-1), seguida pelos dinoflagelados (média: 1,9 x 103 ± 4,7 x 103 cel.L-1); a dominância qualitativa e quantitativa de diatomáceas é uma característica de regiões costeiras.
As duas áreas de estudo apresentaram o mesmo núcleo de espécies principais, variando apenas quanto a algumas espécies consideradas na campanha de 2002 como raras (dinoflagelados em Itaorna e diatomáceas bentônicas em Piraquara de Fora). Dezoito espécies se destacaram em termos de ocorrência e abundância. Deve ser dada atenção especial às espécies do gênero Pseudo-nitzschia (gênero constante e dominante), visto que das 23 espécies descritas até o presente nove são potencialmente tóxicas. Entre os dinoflagelados foram encontradas espécies registradas como formadoras de florações, como P. micans, Scrippsiella sp. e Ceratium hircus.
Embora estudos anteriores tenham caracterizado a região como homogênea em termos hidrológicos e da dinâmica do fitoplâncton, na campanha de 2002 foi detectada heterogeneidade na distribuição espacial das populações, seja de um ponto de coleta para outro, seja considerando cada área isoladamente. As densidades celulares de Piraquara de Fora foram inferiores que às de Itaorna.
Em Itaorna, os gradientes de densidade celular de superfície e de 5 m de profundidade foram análogos e as diferenças em termos de distribuição horizontal eram esperadas, pois a malha amostral incluiu pontos distantes do ponto monitorado (Z3), afastados da linha de costa. Em Piraquara de Fora foi observada homogeneidade em superfície e gradiente a 5 m de profundidade, resultados que alteram (e complementam) a tendência conhecida para a área de estudo.
A abordagem segundo grupos morfofuncionais foi aplicada aos dados das radiais, em caráter preliminar, na tentativa de avaliar a heterogeneidade observada. Foi nítida a estratificação horizontal, tanto em Itaorna quanto em Piraquara de Fora, estabelecida pela relação entre diatomáceas alongadas, mais adaptadas a ambientes com disponibilidade de nutrientes (meso a eutrófico) e maior turbulência, e flagelados (em sua maioria, dinoflagelados), que se adaptam a ambientes com menos nutrientes e verticalmente mais estáveis. Vale destacar que o gradiente horizontal de Piraquara de Fora à superfície foi o inverso do observado a 5 m de profundidade: enquanto na superfície as concentrações de diatomáceas mostraram-se crescentes com o aumento da distância da costa, a 5 m de profundidades as concentrações foram decrescentes.
A variação do fitoplâncton no período de 24 horas foi mais acentuada em Piraquara de Fora do que em Itaorna, especialmente a 5 m de profundidade. Não foi possível associar a variação à oscilação da maré, porém as maiores densidades foram observadas no final do dia, indicando que a luz é um fator controlador do fitoplâncton local.
Estudos futuros devem expandir a malha amostral atual, incluindo duas profundidades e pontos além de R4, de modo a melhor captar a extensão dos efeitos da pluma térmica. Por outro lado, a interpretação de dados de monitoramento, comumente em escala mensal, deve levar em conta a importância de microvariações não detectadas. Nesse sentido, somente uma série temporal de longa escala (anos) poderá minimizar tais diferenças e extrair tendências gerais para o fitoplâncton local.
7.2.3. Fitobentos - topo
As algas marinhas bentônicas, por serem organismos totalmente dependentes da água que as circundam, são consideradas excelentes indicadoras da qualidade da água do local. Algumas espécies podem apresentar sensibilidade a fatores ambientais diversos; em situações extremas, tais fatores podem acarretar o desaparecimento dessas espécies e alterar a estrutura das comunidades onde se encontram.
As espécies de macroalgas podem responder diferentemente aos fatores ambientais. Estas respostas são expressas como alterações nas taxas de fotossíntese, crescimento, produção de elementos de reprodução (esporos e gametas) e produção de metabólitos secundários. Dessa forma, alterações na fisiologia das plantas podem influenciar direta ou indiretamente sua abundância, competitividade e capacidade de ocupar substratos disponíveis.
Por constituírem a base da cadeia alimentar marinha, flutuações no metabolismo das algas, com reflexos em sua biomassa, irão interferir na disponibilidade de alimento para herbívoros, que, por sua vez, poderão comprometer outros elos da cadeia alimentar (Lobban & Harrison 1995).
A utilização de macroalgas em estudos de monitoramento ambiental pode se dar de duas maneiras básicas: estudos in situ e bioensaios, ambos amplamente referidos na literatura, havendo posições a favor e contra cada uma deles (Underwood 1992, 1993). No entanto, é importante destacar que estudos que determinam o grau de tolerância de espécies de algas a diferentes fatores, geralmente estão limitados a testes em laboratório, onde, via de regra, elege-se apenas um fator para ser testado, mantendo-se os demais constantes (Mathews et al. 1982).
Para avaliação das algas marinhas bentônicas foi selecionada a área da Baía da Ilha Grande delimitada pelos seguintes acidentes geográficos: Ponta do Leme (próximo ao Terminal Marítimo Almirante Maximiano, da Petrobras) até Ponta do Mambucaba - no continente, Ponta do Funil até Ponta do Acaiá - na Ilha Grande. Foi priorizada a área de influência direta das usinas, especialmente, a Baía da Ribeira, onde se localiza a descarga dos efluentes das unidades 1 (Angra 1) e 2 (Angra 2) (Figura 59).
O Anexo 7 fornece a listagem das espécies de algas marinhas bentônicas pertencentes às Divisões Rhodophyta, Phaeophyta e Chlorophyta, registradas na literatura para a área de estudo, segundo a nomenclatura e classificação adotadas em Wynne (1998). Não foram considerados os representantes bentônicos das divisões Cyanophyta (cianobactérias) e Chrysophyta (principalmente diatomáceas).
Ao todo, são citadas 242 espécies (50 Chlorophyta, 41 Phaeophyta, 151 Rhodophyta). A ordem mais representativa é a Ceramiales (68 espécies), seguida de Cladophorales e Dictyotales (14 espécies cada).
Os estudos que tratam de algas marinhas bentônicas das proximidades da CNAAA (Unidades 1 e 2), foram classificados, em função de sua abordagem, em: florística (15 referências), taxonomia (1 referência), ecologia (7 referências), toxicologia (1 referência), bioacumulação (5 referências) e química (4 referências). Seus principais resultados foram salientados e comentados. Algumas observações acompanham a apresentação desses estudos.
Ênfase especial foi dada ao estudo de monitoramento ambiental, realizado pela Eletronuclear, por ser o de mais longa duração na área.
Do total de referências analisadas, a maioria é representada por estudos florísticos. Apesar dos estudos florísticos e taxonômicos realizados, é oportuno frisar que o conhecimento acumulado não é satisfatório nem suficiente para representar a flora marinha ficológica da Baía da Ilha Grande, como um todo, considerando sua diversidade de ambientes e a crescente influência antropogênica na área de estudo, como as atividades portuárias, de navegação, e as relacionadas aos crescimentos urbano e turístico.
Figura 59 - Área estudada, com a indicação dos 12 pontos de coleta.
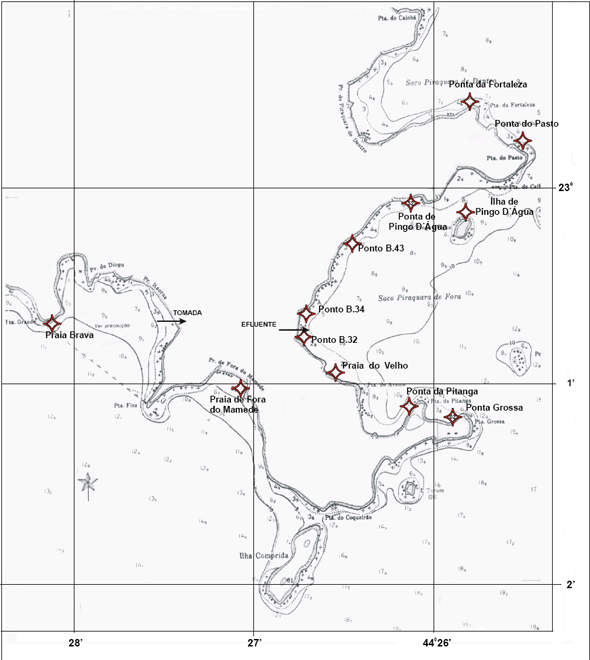
A referência mais antiga às algas marinhas bentônicas da área de estudo data de 1922 (Luetzelburg) e refere-se a um estudo florístico. De lá para cá, apesar de levantamentos florísticos terem sido realizados, houve uma nítida diversificação na abordagem dos estudos, principalmente a partir da década de 80, quando a instalação de Angra 1 motivou estudos de ecologia, toxicologia e bioacumulação.
A Tabela 47 compara a flora da área de estudo com a de outros trechos do litoral sudeste do Brasil. Guardadas as devidas proporções, a flora da área de estudo (251 espécies) pode ser considerada tão rica quanto a flora da região de Arraial do Cabo (Yoneshigue 1985) com 245 espécies. Áreas adjacentes à região de Angra dos Reis, como a Baía de Sepetiba (Pedrini 1980) e o município de Parati (Figueiredo 1989), apresentaram um número inferior de espécies, 180 e 121 espécies, respectivamente. Em todos os estudos, os representantes da Divisão Rhodophyta foram os mais numerosos.
Tabela 47 - Comparação da flora da área de estudo com a de outros trechos do litoral sudeste do Brasil, em relação ao número de espécie e à representatividade dos grupos taxonômicos.
Grupos taxonômicos
Autores-área de estudo |
Divisão Chlorophyta |
Divisão Phaeophyta |
Divisão Rhodophyta |
Total |
| n. spp. |
% |
n. spp. |
% |
n. spp. |
% |
| Yoneshigue-Braga (1970-73): Baía de Guanabara |
27 |
25 |
14 |
13 |
65 |
62 |
106 |
| Yoneshigue (1985): Arraial do Cabo |
43 |
17 |
47 |
19 |
155 |
64 |
245 |
| Pedrini (1980): Baía de Sepetiba |
37 |
20 |
27 |
15 |
116 |
65 |
180 |
| Figueiredo (1989): Município de Parati |
23 |
18 |
28 |
23 |
70 |
59 |
121 |
| Região de influência de Angra 3 |
53 |
23 |
42 |
17 |
156 |
60 |
251 |
Dentre os estudos florísticos na área de estudo, ressaltamos a importância do relatório pré-monitoramento, realizado pela equipe de especialistas da Universidade Federal do Rio de Janeiro, sob coordenação do Prof Gilberto J. P. Mitchell (FUJB 1980/81), onde vários pontos de coleta, nas proximidades do ponto de lançamento do efluente térmico da CNAAA, foram amostrados em épocas diferentes, por um ano, englobando as 3 Divisões de macroalgas.
Estudo semelhante foi realizado pela equipe de Alexandre de G. Pedrini, do então Instituto de Radioproteção e Dosimetria/ Cnen, que também levantou a flora local (Pedrini & Pereira, 1983). Esse estudo também deve ser considerado como de pré-monitoramento, pois as usinas ainda não estavam em pleno funcionamento na época das coletas.
Para o período após o início de funcionamento das usinas, não contamos com estudos florísticos comparáveis aos de FUJB (1980/81) e Pedrini& Pereira (1983). Os únicos estudos florísticos realizado nas proximidades da saída do efluente é o de Maria Teresa M. de Széchy (Széchy 1996, Széchy & Paula 2000), que acompanhou as comunidades de Sargassum em diferentes épocas do ano, por cerca de um ano. Apesar de ter se restringido à região sublitorânea rasa, onde espécies de Sargassum estavam presentes, os resultados em termos de listagem de espécies são interessantes para uma análise temporal, pois seus três pontos de coleta coincidiram com pontos também selecionados no estudo de pré-monitoramento. A composição de espécies de feofíceas presentes em um único ponto (32B1) próximo ao lançamento do efluente da CNAAA ao longo de 12 anos é analisada por Costa (1996), com base nos dados do monitoramento ambiental de Angra 1 e 2.
A flora marinha bentônica de profundidade da área de estudo permanece desconhecida, apesar do estudo recente de Horta (2000) para algumas ilhas da Baía da Ilha Grande.
Em floras marinhas tropicais, as rodofíceas são as macroalgas melhor representadas, em termos de número de espécies (Lüning 1990). Daí, seu papel central na diversidade específica das comunidades bentônicas. Sendo assim, vemos como fundamental o estudo desse grupo na área sob influência das usinas, além das feofíceas, como vem sendo feito.
O estudo da flora marinha bentônica, como um todo, também se justifica por outro aspecto, recentemente valorizado por agências de pesquisa institucionais, que é o problema de introdução de espécies no meio marinho, algumas das quais podem se tornar invasoras e indesejáveis. Exemplos conhecidos são os da alga verde Caulerpa taxifolia e da alga parda Sargassum muticum (Critchley et al. 1983; Meinesz et al. 1993). A definição de quais espécies é exótica ou não para a região depende diretamente do conhecimento pretérito da flora, como um todo.
Outra justificativa para o estudo da flora marinha, como um todo, pauta-se na necessidade de conhecimento do processo de colonização e estruturação das comunidades marinhas bentônicas. De longa data, é conhecido que esses processos ocorrem a partir da interação entre as diferentes espécies presentes (Dayton 1971). Com relação as macroalgas, a capacidade de competição depende de características morfológicas e fisiológicas das espécies, ou seja de seu tipo morfo-funcional (Steneck & Dethier 1994), e não de sua identidade taxonômica. Nesse contexto, clorofíceas, feofíceas e rodofícias podem ser equiparadas quanto à capacidade de colonização e domínio em comunidades marinhas bentônicas. Para exemplificar o papel das rodofíceas na estruturação de comunidades da área de estudo, podemos nos reportar a estudo mais recente, na Enseada de Monsuaba, contígua à Baía da Ribeira, também na Baía da Ilha Grande (Falcão 1996). Esse estudo, ao acompanhar, por cerca de um ano, o comportamento de comunidades bentônicas da região entre-marés, através dos dados de cobertura, sob a influência ou não de distúrbios provocados, mostra que as algas vermelhas filamentosas, pertencentes à ordem Nemalionales (Asparagopsis taxiformis), estão entre as mais comuns na região.
Acompanhamentos temporais de aspectos qualitativos e quantitativos das comunidades fitobentônicas foram realizados antes e depois da instalação da usina (FUJB 1980/81; Pedrini et al. 1994b; Falcão 1996; Széchy 1996). Apesar de não conclusivos, esses estudos apontam para dois padrões de comportamento temporal divergentes: comunidades não submetidas à massa d’água aquecida pela usina tendem a tornar-se mais exuberantes em termos de cobertura da vegetação e/ou biomassa no período primavera-verão; comunidades submetidas à essa massa d’água aquecida tendem a apresentar maior abundância de macroalgas no período outono-inverno, especialmente no caso de comunidades onde Sargassum ocorre. No entanto, não é conhecida a capacidade de restabelecimento das espécies de algas que existiam na Baía da Ribeira antes da instalação das usinas nucleares, e que vêm diminuindo em termos de freqüência e de abundância, frente aos distúrbios contínuos a elas impostos há mais de 20 anos. Essa situação é particularmente crítica no caso dos bancos de Sargassum, por seu reconhecido papel como habitat para muitas outras espécies vegetais e animais (Norton & Benson 1983; Edgar 1983; Széchy & Paula 1997; Széchy et al. 2001).
Para o estudo de fitobentos, foram selecionados 12 pontos de coleta (Figura 60), oito dos quais situados no Saco Piraquara de Fora, área de lançamento dos efluentes das usinas Angra 1 e Angra 2 e, futuramente, Angra 3, sendo três desses pontos no sentido da Ponta Grossa e quatro, no sentido da Ilha de Pingo d’Água. Os quatro pontos restantes foram localizados em áreas consideradas como controle, adjacentes ao Saco Piraquara de Fora, posicionadas mais ao norte (dois pontos no Saco Piraquara de Dentro) e ao sul (dois pontos adjacentes à Itaorna). Sempre que possível, os pontos de coleta foram posicionados em locais que fazem ou já fizeram parte do programa da Eletronuclear para monitoramento dos efluentes da refrigeração.
O estudo concentrou-se nas comunidades de substrato rochoso da região sublitorânea rasa (até cerca de 5 m de profundidade), porém incluiu a franja da região sublitorânea, ou seja, a parte do costão que fica emersa durante as marés de sizígia. Coleta de material e as observações foram feitas em apnéia e por meio de mergulho autônomo.
A coleta foi feita nos dias 3, 4, 5, 6 e 30 de maio de 2002, sendo cada ponto de amostragem visitado pela equipe apenas uma vez, ocasião em que foram medidas a salinidade e a temperatura da água do mar. Em cada ponto foram realizadas amostragem não destrutiva, amostragem destrutiva em quadrados e amostragem destrutiva por busca.
Para a amostragem não destrutiva, foram delimitados em cada ponto três transetos de 5 m de extensão, divididos em 50 pontos equidistantes (10 cm). Os transetos foram posicionados perpendicularmente à linha d’água e paralelamente entre si, distantes um do outro cerca de 1 m, com início na franja da região sublitorânea (marca 1 do transeto), onde se notou a ocorrência de macroalgas. Os organismos presentes em cada um dos 50 dos transetos foram registrados e identificados em nível específico. Quando a identificação segura em nível específico in situ não foi possível, os organismos foram identificados em categorias taxonômicas superiores ou classificados de acordo com seu grupo morfofuncional segundo os critérios de Steneck & Dethier (1994) e Széchy (1996). Essa abordagem forneceu a distribuição vertical dos organismos no costão rochoso, dados com base nos quais foi estimada a dominância relativa no estrato superior, bem como propostos padrões de zoneamento para a área de estudo.
Para a amostragem destrutiva em quadrados foram lançados ao acaso três quadrados de 0,25 m de lado em dois níveis do costão rochoso, definidos em função da temperatura da água do mar e da profundidade:
-
nível raso (n=3) ® temperatura da água do mar mais elevada; profundidade entre 0,5 e 1,5m (foi evitada a franja da região sublitorânea);
-
nível fundo (n=3) ® temperatura da água do mar nitidamente mais fria; profundidade entre 2,0 e 4,0m.
A área interna dos quadrados foi raspada com o auxílio de espátulas metálicas e o material acondicionado em sacolas de náilon etiquetadas, constituindo as amostras. Posteriormente, o material foi acondicionado em sacos e/ou potes plásticos e fixado em solução de formaldeído a 4%. Dessa forma, o tamanho amostral da variável biomassa foi (n=6) para todos os pontos de coleta, exceto o situado mais próximo do local de descarga da água de resfriamento, onde não foi notada diferença de temperatura da água do mar entre a superfície e o fundo, para o qual foi adotado (n=3). Essa amostragem forneceu a estimativa de biomassa (aspecto quantitativo) das macroalgas e a freqüência relativa (aspecto quantitativo) das espécies, em ambos os casos por ponto de coleta e por nível no costão rochoso, bem como complementou a listagem das espécies (aspecto qualitativo). Espécies registradas em 6 ou 5 dos quadrados amostrados de um ponto de coleta foram consideradas como muito freqüentes nesse ponto de coleta.
Para a amostragem por busca, costões, matacões e blocos rochosos situados desde a franja da região sublitorânea até a profundidade de ocorrência (cerca de 5 m), numa extensão horizontal de aproximadamente 50 m, foram minuciosamente inspecionados para a coleta da maior diversidade possível de macroalgas. Foram coletados exemplares epilíticos, epifíticos e epizóicos, exceto os fortemente aderidos ao substrato, ou seja, os incrustantes. O material coletado foi transportado para a embarcação em sacolas de náilon, sendo logo armazenado em sacos plásticos etiquetados e fixado em solução de água do mar e formaldeído a 4%. Essa abordagem forneceu a composição específica das macroalgas (aspecto qualitativo) dos pontos de coleta e, complementada pela listagem de espécies oriunda da triagem das amostras de biomassa, forneceu também a riqueza de espécies de macroalgas (aspecto quantitativo) dos pontos de coleta.
Em laboratório, as amostras provenientes da amostragem destrutiva em quadrados foram limpas, triadas e identificadas em nível infragenérico (quando possível); em alguns casos, macroalgas de mesmo tipo morfofuncional foram pesadas juntas. O peso seco das macroalgas triadas foi obtido após secagem em estufa a 70oC até atingirem peso constante. O material foi pesado em balança digital, com precisão de 0,01 grama.
Figura 60 - Fitobentos - Pontos de coleta
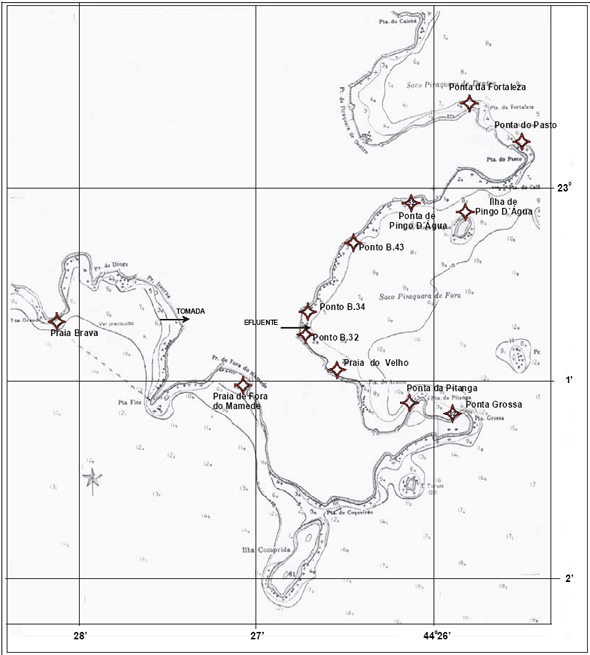
O material proveniente da amostragem destrutiva por busca foi triado sob microscópio estereoscópico e identificado em nível infragenérico, adotando-se Joly (1965) como publicação básica para a identificação taxonômica, complementada, quando necessário, por publicações mais recentes e especializadas, principalmente da literatura nacional. A taxonomia das espécies seguiu a nomenclatura revisada proposta por Wynne (1998).
Material dos gêneros Cladophora (Chlorophyta) e Acrochaetium (Rhodophyta) foi enviado a especialistas para identificação, respectivamente Lísia Gestinari, da Universidade Federal de Pernambuco, e Joel Campos de Paula, da Universidade do Estado do Rio de Janeiro. Parte do material de Laurencia (Rhodophyta) teve sua identificação confirmada pela especialista Mutue Toyota Fujii, do Instituto de Botânica de São Paulo. Nos casos de insuficiência de material (quantidade coletada pequena), material fragmentado, material não fértil e material requerendo técnicas especiais para identificação taxonômica segura, as macroalgas foram identificadas apenas em nível genérico e registradas como spp. As cianobactérias, apesar de registradas, não foram tratadas taxonomicamente.
Foram calculados a média aritmética, o desvio padrão e o coeficiente de variação (coef var= desvio padrão/média x 100) do peso seco das espécies, por ponto de coleta e por nível no costão rochoso.
7.2.3.1. Caracterização da Comunidade Fitobentônica - topo
A afinidade florística da área de estudo com áreas geográficas próximas, do litoral sul do estado do Rio de Janeiro e norte do estado de São Paulo, é facilmente reconhecida pelo elevado número de espécies em comum (Joly, 1965; Oliveira Filho, 1977; Figueiredo, 1989; Falcão et al., 1992).
De fato, este trecho do litoral brasileiro é considerado como pertencente a uma única região ficogeográfica, por diferentes autores. Mitchell et al. (1979), revisando as clorofíceas bentônicas do litoral do estado do Rio de Janeiro, classificam o trecho correspondente aos municípios de Angra dos Reis e Parati como uma única área, ressaltando que sua flora apresenta componentes comuns ao litoral norte do estado de São Paulo e à Baía de Sepetiba. Oliveira Filho (1977) posiciona a área de estudo na Zona Sudeste, caracterizada por uma flora marinha bem desenvolvida. Horta (2000) posiciona a área de estudo na Região Temperada Quente, que apresenta seu limite norte ao norte do estado do Rio de Janeiro e o limite sul, na região de Rio Grande, no estado do Rio Grande do Sul. Em escala mais ampla, Lüning (1990) inclui a região sul do estado do Rio de Janeiro, onde insere-se a área de estudo, bem como o restante do litoral brasileiro ao sul, em uma região biogeográfica marinha ampla, denominada região temperada quente do Hemisfério Sul, reconhecida por mudanças drásticas da flora e fauna litorâneas e delimitada pelas isotermas da água de superfície: 25oC (no verão ) e 20oC (no inverno).
Considerando-se os índices de Feldmann (1937) (relação entre o número de espécies de Rhodophyta sobre o de Phaeophyta) e de Cheney (1977) (relação entre o número de espécies de Rhodophyta e de Chlorophyta sobre o de Phaeophyta), os valores encontrados para a flora da área de estudo (R/P = 3,1; R+C/ P = 4,4) coincidiram com os propostos para regiões subtropicais/ temperadas quentes (R/P < 4,0; R+C/ P < 6,0), como o esperado em função do seu posicionamento geográfico. Apesar da temperatura da água do mar da área de estudo, especialmente a do Saco Piraquara de Fora, ter sofrido considerável aumento após o lançamento do efluente térmico de Angra 1 e 2, esta mudança não parece ter causado, até o presente momento, a exclusão ou a substituição de parcela significativa de espécies da flora marinha, de um modo geral. Isto não se aplica quando os pontos de coleta são considerados isoladamente, e quando são analisados aspectos quantitativos e de distribuição vertical das espécies.
Das 101 espécies encontradas no levantamento realizado durante a fase “pré-operacional” de Angra 1, 73 espécies tornaram a ser coletadas (72,3%). Deve ser levado em consideração que a listagem de espécies de macroalgas ora apresentada corresponde a um esforço amostral menor do que o do “pré-operacional”, tanto em relação ao número de pontos de coleta como em relação ao número de coletas efetuadas por ponto. Por outro lado, deve ser ressaltado que a presente listagem de espécies é resultado da identificação de material por um maior número de especialistas, o que certamente possibilitou o aprimoramento da parte taxonômica.
Com base no levantamento dos dados pretéritos para a área de influência da Central Nuclear Almirante Álvaro Alberto, 21 espécies citadas neste estudo correspondem a novas ocorrências para a área. Dessas novas ocorrências, 13 são da Divisão Rhodophyta, 1 de Phaeophyta e 6 de Chlorophyta, além de uma espécie de alga verde (Pseudoderbesia arbuscula ?), cuja identificação definitiva está na dependência da comparação com o material tipo, podendo tratar-se de uma referência nova para o litoral do Brasil.
As ocorrências novas para a área de estudo podem ser interpretadas das seguintes formas:
-
Espécies criptogênicas, que são espécies raras, de ocorrência esporádica e ambientes mais restritos; algumas destas espécies, apesar de citadas para o litoral norte do Estado de São Paulo (Joly, 1965), estão sendo somente agora citadas para a área de estudo, talvez pela dificuldade de coleta e triagem, por causa de suas pequenas dimensões;
-
Espécies introduzidas recentemente, através de mecanismos diversos, favorecidas talvez pelas alterações na estrutura das comunidades dos costões rochosos.
Dentre as espécies anteriormente citadas para área de influência da CNAAA por diferentes autores, 138 espécies (94 Rhodophyta + 19 Phaeophyta + 25 Chlorophyta) não foram identificadas no presente estudo. A princípio, esta diferença impressiona, mas, na verdade, ela está superestimada. Deve-se levar em consideração que nossos resultados refletem uma amostragem menor, em termos de área e de escala temporal, do que os dados compilados na listagem de dados pretéritos. Dentre as espécies não identificadas por nós, incluem-se grupos que devem ser revistos, a saber: 1) aqueles com problemas taxonômicos e que necessitam de revisão (Exemplo: Bryopsis spp., Derbesia spp., Ceramium spp., Dasya spp., Gracilaria spp., Laurencia spp.); 2) aqueles com problemas nomenclaturais e de sinonimia (Exemplo: Derbesia marina, Fosliella chamaedoris); 3) aqueles de coleta e/ou triagem mais difíceis, pelas suas dimensões reduzidas ou pelo seu hábito incrustante (Exemplo: Antithamnion villosum, Diplothamnion tetrastichum, Dipterosiphonia dendritica, Dohrniella antillarum, Gymnothamnion elegans, Myrionema strangulans, Peyssonnelia spp., Lithothamnion spp.); 4) aqueles de habitats mais restritos, não inspecionados durante este levantamento, que priorizou a região sublitorânea de substratos rochosos e locais protegidos do embate das ondas (Exemplos: Avrainvillea elliottii, Bostrychia spp., Chnoospora minima, Gayralia oxysperma, Hincksia breviarticulata, Ophidocladus herposiphonioides, Porphyra spp.); 5) aqueles de ocorrência rara (Exemplo: Crouania attenuata, Nemacystus howei); 6) aqueles de ocorrência duvidosa (Exemplo: Valonia aegagropila, Valonia utricularis). Finalmente, devemos levantar a hipótese do desaparecimento de algumas espécies anteriormente citadas, em função de fatores limitantes ao seu crescimento e reprodução, resultantes das mudanças da massa d’água na área de estudo. Dentre estes fatores, a temperatura da água do mar é de indiscutível importância e será discutida mais adiante (vide item 3.1.9).
Foram identificadas, ao todo, 124 espécies de macroalgas, sendo 29 Chorophyta, 23 Phaeophyta e 72 Rhodophyta. A maior representatividade, em termos de número de espécies, foi encontrada nas ordens Ceramiales (36 espécies), Dictyotales (11 espécies) e Cladophorales (12 espécies).
A sinopse dos táxons encontrados durante o levantamento encontram-se no Anexo 8.
A lista os táxons identificados, por Divisão, nos 12 pontos de coleta é apresentada Anexo 9. Esta listagem engloba o material oriundo da coleta por “busca” em diferentes micro-ambientes da região sublitorânea do ponto de coleta, bem como o material oriundo da triagem dos quadrados raspados no nível raso (n=3) e no nível fundo (n=3). A partir destes dados, foram obtidas a composição e a riqueza específicas de cada ponto de coleta, bem como a freqüência das espécies.
O ponto de coleta com maior número de espécies foi a Ponta da Fortaleza, seguido da Ponta do Pasto, ambos no Saco Piraquara de Dentro. Nestes pontos, também foram encontrados os maiores números de espécies de Phaeophyta, grupo considerado bom indicador da qualidade do ambiente marinho (Borowiztka, 1972; Berchez & Oliveira, 1992). O número de espécies de Phaeophyta diminuiu drasticamente do Saco Piraquara de Dentro (área controle) para o local de lançamento do efluente térmico, no Saco Piraquara de Fora (Ponto B.32), onde foram encontradas apenas 2 espécies do mesmo gênero (Sphacelaria).
É interessante ressaltar que estas espécies só foram encontradas férteis por propágulos - estruturas de reprodução vegetativa. Em Sphacelaria, a produção de propágulos, esporângios ou gametângios está vinculada à temperatura da água do mar e ao fotoperído; temperaturas elevadas inibem a diferenciação de gametas e, assim, interferem na alternância de gerações (van den Hoek & Flinterman, 1968; Colijn & van den Hoek, 1971).
Os resultados dos levantamentos realizados estão de acordo com Verlaque (1976) e Verlaque et al. (1981). No Mediterrâneo, também foi notada a diminuição do número de espécies de macroalgas em áreas próximas ao lançamento do efluente térmico. Na área de estudo, cerca de 21% das espécies tiveram ocorrência em mais de 75% dos pontos de coleta, ou seja, 26 espécies foram muito freqüentes na área de estudo (Tabela 48).
Tabela 48- Espécies muito freqüentes na Área de Estudo
| Phaeophyta |
Rhodophyta |
| Dictyopteris delicatula |
Acanthophora spicifera |
Gelidopsis variabilis |
| Dictyota cervicornis |
Amphiroa fragilissima |
Gelidium pusillum |
| Padina gymnospora |
Asparagopsis taxiformis |
Hypnea musciformis |
| Sargassum vulgare |
Ceramium flaccidum |
Hypnea spinella |
| Sphacelaria rigidula |
Champia parvula |
Jania adhaerens |
| |
Champia vieillardii |
Laurencia papillosa |
| |
Chondracanthus acicularis |
Polysiphonia ferulacea |
| |
Dasya spp. |
Pneophyllum fragile |
| |
Erythrotrichia carnea |
Stylonema alsidii |
| |
Galaxaura marginata |
Wrangelia argus |
| |
Gelidiopsis planicaulis |
|
S = 26 |
Por outro lado, cerca de 26% das espécies foram encontradas apenas em um ponto de coleta (Tabela 49).
Tabela 49 - Espécies encontradas em apenas um ponto na Área de Estudo
| Chlorophyta |
Phaeophyta |
Rhodophyta |
| Acicularia schenckii |
A. rhodochortonoides |
Acrochaetium flexuosum |
| Bryopsis hypnoides |
Bachelotia antillarum |
Acrochaetium hallandicum |
| Chaetomorpha aerea |
Dictyota crenulata |
Antithamnion lherminieri |
| Chaetomorpha nodosa |
Feldmannia irregularis |
Bryothamnion seaforthii |
| Cladophora coelothrix |
Lobophora variegata |
Ceramium tenerrimum |
| Cladophora corallicola |
Rosenvingea intricata |
Chondria polyhriza |
| Cladophora prolifera |
Spatoglossum schroederi |
Gelidiella trinitatensis |
| Codium decorticatum |
Sphacelaria brachygonia |
Gelidium crinale |
| Codium intertextum |
|
Grateloupia cuneifolia |
| Ulva lactuca |
|
Gymnogongrus griffithsiae |
| |
|
Halymenia bermudensis |
| |
|
Laurencia scoparia |
| |
|
Lomentaria corallicola |
| |
|
Polysiphonia tepida |
S = 32 |
Excluindo Erythrotrichia carnea e Stylonema alsidii, espécies notadas apenas sob microscópio, todas as demais espécies consideradas muito freqüentes no presente estudo foram citadas anteriormente para o Saco Piraquara de Fora (FUJB, 1980/1981; Pedrini et al. 1994a; Széchy, 1996; Széchy & Paula, 2000a). Dentre estas espécies, Dictyopteris delicatula, Dictyota cervicornis, Padina gymnospora, Amphiroa fragilissima, Asparagopsis taxiformis, Champia vieillardii, Gelidiopsis planicaulis, Gelidiopsis variabilis, Hypnea musciformis, Hypnea spinella, Jania adhaerens e Pneophyllum fragile foram consideradas como características dos bancos de Sargassum da área de Angra dos Reis, devido à sua alta freqüência (Széchy, 1996). Este conjunto de espécies deve ser utilizado em monitoramentos futuros para acompanhar as mudanças na estrutura das comunidades fitobentônicas da região sublitorânea. O desaparecimento de espécies deste conjunto pode ser interpretado como reflexo provável de alterações no ambiente.
O número de espécies por ponto de coleta e por nível do costão rochoso pode ser sumarizado na Tabela 50.
Tabela 50 - Numero de espécies por ponto de coleta e por nível do costão rochoso na Área de Estudo
Pontos de coleta |
Número de espécies |
| Raso |
Fundo |
Total |
| Praia Brava |
30 |
29 |
49 |
| Praia de Fora do Mamede |
34 |
31 |
50 |
| Ponta Grossa |
15 |
27 |
47 |
| Ponta da Pitanga |
22 |
23 |
55 |
| Praia do Velho |
21 |
29 |
51 |
| B. 32 (descarga) |
8 |
- |
22 |
| B. 34 |
21 |
27 |
46 |
| B. 43 |
23 |
40 |
47 |
| Ponta de Pingo d’Água |
19 |
26 |
60 |
| Ilha de Pingo d’Água |
17 |
28 |
45 |
| Ponta do Pasto |
22 |
28 |
65 |
| Ponta da Fortaleza |
34 |
34 |
71 |
A maior riqueza específica foi encontrada nos dois pontos de coleta do Saco Piraquara de Dentro: Ponta da Fortaleza (71 espécies) e Ponta do Pasto (65 espécies). Em relação ao nível do costão, o maior número de espécies foi encontrado no fundo (Ponto B.43, com 40 espécies), enquanto o menor foi encontrado no raso (Ponto B.32, com 8 espécies). É interessante notar que em todos os pontos de coleta do Saco Piraquara de Fora, o maior número de espécies foi encontrado no nível fundo.
Comparando-se o número de espécies do nível fundo da Ponta da Fortaleza, Ponta de Pingo d’Água e Ponta da Pitanga, com os dados fornecidos por Széchy (1996) e Széchy & Paula (2000a), para bancos de Sargassum dos mesmos pontos de coleta, estudados anteriormente, podemos constatar uma diminuição do número de espécies nos pontos de coleta do Saco Piraquara de Fora. A Tabela 51 a seguir compara o número de espécies de macroalgas no nível fundo (banco de Sargassum), em pontos de coleta da Baía da Ribeira, em 1991-1992 e em 2002.
Tabela 51 - Comparação entre número de espécies no nível fundo na Área de Estudo
Pontos de coleta |
Número de espécies |
| Presente estudo |
Széchy (1996) |
Széchy & Paula (2000a) |
| Ponta da Fortaleza |
34 |
25- 44 |
15- 31/m2 |
| Ponta de Pingo d’Água |
26 |
37 - 43 |
22- 32/m2 |
| Ponta da Pitanga |
23 |
43- 48 |
30- 34/m2 |
O levantamento taxonômico sugere a tendência da estrutura das comunidades bentônicas da área de estudo, em relação à riqueza de espécies de macroalgas: diminuição do número de espécies da área controle em direção ao ponto de descarga dos efluentes (B. 32); simultaneamente, a diminuição do número de espécies no nível “raso” do costão, na área sob influência direta destes efluentes (Saco Piraquara de Fora).
Esta tendência deve ser analisada com base em dois parâmetros, que refletem gradientes de temperatura da água do mar, entre outros possíveis fatores não considerados. O primeiro destes parâmetros é a distância do costão rochoso à descarga dos efluentes, o que deve ser avaliado em conjunto com o comportamento da circulação da massa d’água superficial aquecida. O segundo parâmetro é a profundidade do costão, o que deve ser avaliado em conjunto com aspectos topográficos, que possam interferir na mistura da camada de água superficial aquecida com a camada de água do fundo.
Dustribuição vertical dos organismos
O Anexo 10 apresenta quadros demonstrativos com os organismos observados nos 12 pontos de coleta, ao longo dos 3 transetos lançados perpendicularmente à linha d’água, a partir da franja da região sublitorânea. A numeração corresponde à posição do organismo no transeto (1=10cm), a partir da franja da região sublitorânea até 5m de extensão vertical. Os organismos foram identificados, no mar, em nível específico, quando possível. Aqueles com dificuldade de identificação taxonômica, em função de suas dimensões ou quaisquer outros motivos, foram classificados em categorias taxonômicas mais elevadas ou de acordo com seu tipo morfo-funcional.
A partir dos dados obtidos com o uso dos transetos, foi possível distinguir diferentes padrões de zonação para a área de estudo, denominados de Tipos 1, 2, 3, 4 e 5.
A Figura 61 mostra, de modo esquemático, os cinco tipos de padrão de zonação. Cada padrão pode ser caracterizado pela seqüência de faixas de organismos dominantes, em termos de cobertura do estrato superior, a partir da franja da região sublitorânea em direção ao fundo de areia. A altura das faixas corresponde à área relativa ocupada pelos organismos referidos, ao longo do costão. É importante ressaltar que, na maioria dos pontos de coleta, os transetos não cobriram toda a extensão vertical do substrato rochoso disponível. Portanto, os padrões estabelecidos restringem-se à parte mais rasa do costão, representando uma extensão de 5m. Assim, a profundidade máxima analisada por ponto de coleta variou, dependendo da inclinação do costão rochoso.
Figura 61 - Esquema dos padrões de zonação definidos para as comunidades bentônicas da área sob influência das Unidades 1 e 2.
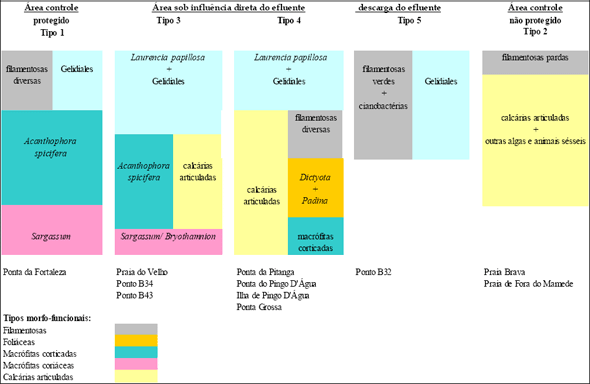
Os padrões de zonação foram agrupados de acordo com a distância da área de descarga do efluente das Usinas Angra 1 e 2, a saber:
Tipo 1: definido para costões protegidos do embate das ondas
Saco Piraquara de Dentro - Ponta da Fortaleza
Tipo 2: definido para costões não protegidos do embate direto das ondas
Ponta do Pasto
Praia Brava
Praia de Fora do Mamede
Área sob influência direta do efluente: Saco Piraquara de Fora
Tipo 3:
Praia do Velho (lado em direção à Ponta Grossa)
B.34 (lado em direção à Ilha de Pingo d’Água)
B.43 (lado em direção à Ilha de Pingo d’Água)
Tipo 4:
Ponta Grossa
Ponta da Pitanga (lado em direção à Ponta Grossa)
Ponta de Pingo d’Água (lado em direção à Ilha de Pingo d’Água)
Ilha de Pingo d’Água
Pode-se concluir que a comunidade bentônica da área de estudo mostrou diferenças expressivas quanto à distribuição vertical e cobertura dos organismos no costão, se comparados os 12 pontos de coleta. Estas diferenças foram traduzidas na multiplicidade de padrões de zonação e na variação do grau de heterogeneidade dos costões quanto ao aspecto fisionômico.
Os padrões de zonação estabelecidos podem ser relacionados à distância do ponto de coleta em relação à descarga do efluente e também às próprias características do costão rochoso, especialmente quanto à exposição ao embate das ondas. Pequenas variações em relação aos padrões estabelecidos são esperadas e podem estar relacionadas a outros fatores abióticos e bióticos e a variações temporais, que fogem do escopo deste estudo.
Foi observado que, em locais protegidos da área controle, o costão apresentava nitidamente 3 faixas (Tipo 1), sendo as mais conspícuas a de Acanthophora spicifera e, abaixo, a de Sargassum spp., macroalgas bastante comuns na Baía da Ilha Grande (Falcão, 1996). Em locais não protegidos da área controle (Tipo 2), exemplares de Acanthophora spicifera e Sargassum spp. também foram registrados, porém sem constituir faixa, por não serem dominantes. Nesses locais, o costão apresentava 2 faixas, sendo a de calcárias articuladas (Corallinaceae) bastante extensa e heterogênea, devido à presença de outras macroalgas e de animais sésseis, crescendo em manchas entre as almofadas de calcárias articuladas.
Nas comunidades bentônicas da área sob influência direta da descarga dos efluentes (Saco Piraquara de Fora), calcárias articuladas aumentaram sua importância quanto à cobertura do estrato superior (Tipos 3 e 4), quando comparadas às comunidades de Tipo 1, enquanto que Acanthophora spicifera e Sargassum spp., diminuíram de importância. No Tipo 3, Acanthophora spicifera crescia junto com calcárias articuladas; Sargassum, em muitos casos, foi exclusivo das rochas situadas mais no fundo, onde a temperatura da água do mar era menor, formando faixas mais estreitas e heterogêneas. No Tipo 4, as calcárias articuladas passaram a ser dominantes; Acanthophora spicifera e Sargassum spp., quando presentes, cresciam isoladamente ou em pequenas manchas, juntamente com macroalgas de outros tipos morfo-funcionais.
Na descarga do efluente, a comunidade bentônica mostrou-se bastante homogênea quanto ao aspecto fisionômico (Tipo 5), sendo definida apenas uma faixa, com domínio de algas de morfologia bastante simples, pertencentes ao grupo morfo-funcional das filamentosas, incluindo verdes e cianobactérias. As características faixas de Acanthophora spicifera e de Sargassum spp. não foram observadas. Macrófitas corticadas também foram expressivas, sendo representadas por Gelidiales.
Os resultados do presente estudo estão de acordo com Verlaque (1976) e Verlaque et al. (1981). Esses autores, em estudo similar realizado no Mediterrâneo, a respeito dos efeitos de efluente térmico sobre o fitobentos marinhos, também observaram mudanças na estrutura da comunidade, em função da distância da descarga do efluente térmico e em função da profundidade. Para o local de descarga do efluente, os autores citam a abundância de macroalgas pequenas e em tufos, de ciclos de vida curtos, e a ausência de macroalgas eretas maiores. Dentre estas espécies, ressaltam a importância de representantes de Cladophora e de Gelidiales, além de cianobactérias filamentosas, como o encontrado na área de estudo. Também descrevem o aspecto “monótono e monocromático, dando a ilusão da ausência de fitobentos na área impactada.
É oportuno salientar que bancos densos de Sargassum, descritos para vários pontos de coleta da área de estudo, antes do início de funcionamento da Usina Angra 1 (FUJB, 1980/1981),foram ainda encontrados em 1991/1992 (Széchy, 1996), em três dos pontos de coleta estudados agora: Ponta da Fortaleza, Ponta da Pitanga e Ponta de Pingo d’Água. A autora fornece dados de cobertura nos bancos, durante um ano; a cobertura de Sargassum nesses bancos variou de 28% a 96%. Além disso, espécies de Sargassum correspondiam às plantas mais desenvolvidas em termos de altura, sendo reconhecidas como clímax das comunidades bentônicas de substrato duro (Széchy, 1996). A superioridade competitiva de Sargassum, em ambientes sem distúrbios, é ressaltada na literatura, inclusive para espécies do litoral brasileiro (Eston & Bussab, 1990).
Caracterização dos Pontos de Coleta
Coordenadas: 23º01’0.3”S e 44º29’10.6” W
Costão exposto ao embate das ondas, de inclinação acentuada.
Profundidade: 7,6m
Temperatura à superfície:26,5 ºC
Temperatura no fundo:26 ºC
Salinidade: 33
Foram identificadas 49 espécies de algas bentônicas, sendo 05 Chlorophyta, 09 Phaeophyta e 35 Rhodophyta. Destas, as mais freqüentes foram Dictyopteris delicatula, Sargassum filipendula, Aglaothamnion cf. tenuissimum, Amphiroa fragilissima, Gelidiopsis variabilis, Hypnea musciformis, Jania adhaerens e as epífitas Asparagopsis taxiformis- fase tetrasporofítica, Ceramium flaccidum e Pneophyllum fragile, que ocorreram em unidades amostrais do raso e do fundo. Caulerpella ambigua, Amphiroa beauvoisii e Dasya spp.ocorreram em todas as unidades amostrais do raso, enquanto Champia vieillardii e Chondrachanthus acicularis ocorreram em todas as unidades amostrais do fundo. Cianobactérias filamentosas (Oscillatoriaceae) foram observadas, mas não identificadas.
Em termos de cobertura do estrato superior, foram identificadas três faixas ao longo dos transetos: superior (0-0,4m), com domínio de algas filamentosas; intermediária (0,5-4,2m), com domínio de calcárias articuladas (Corallinaceae); inferior (4,3-5,0m), com domínio de macrófitas corticadas, como Gelidiopsis spp. e Hypnea spp., e de macrófitas coriáceas, no caso Sargassum spp., no entanto, sem formar populações densas. Manchas de diferentes organismos mesclavam-se dentro da faixa de calcárias articuladas: Laurencia spp. mais acima (0,7-2,0m), Gelidiales (1,0-3,2m), Sargassum spp. (1,1-3,6m), Dictyota spp. (0,7-3,5m) e Asparagopsis taxiformis mais abaixo (2,9-4,0m). Ouriços pretos (Echinometra lucunter)ocorreram em toda a extensão vertical do costão, em grande quantidade.
A área amostral deste ponto de coleta mostrou-se bastante heterogênea quanto ao aspecto fisionômico, se comparada às demais.
Em termos de biomassa, as espécies mais representativas foram Gelidium pusillum (86,72g/m2- valor médio, no raso), Sargassum filipendula (41,28g/m2- valor médio, no fundo) e Asparagopsis taxiformis - fase tetrasporofítica (18,51g/m2- valor médio, no fundo). As calcárias articuladas (Corallinaceae) foram mais expressivas no raso (valor médio= 99,73g/m2); Tricleocarpa fragilis mostrou biomassa pouco expressiva no fundo (2,83 g/m2).
Coordenadas: 23º01’10.7”S e 44º27’2,4” W
Costão moderadamente exposto ao embate das ondas, de inclinação acentuada.
Profundidade: 7,2m
Temperatura à superfície:27 ºC
Temperatura no fundo:26 ºC
Salinidade: 36
Foram identificadas 50 espécies de algas bentônicas, sendo 06 Chlorophyta, 07 Phaeophyta e 37 Rhodophyta. Destas, as mais freqüentes foram Dictyopteris delicatula Dictyota cervicornis, Sargassum filipendula, Amphiroa fragilissima, Gelidiopsis variabilis, Laurencia filiformis, Peyssonnelia spp. e a epífita Asparagopsis taxiformis- fasetetrasporofítica, que ocorreram em unidades amostrais do raso e do fundo. Sargassum vulgare, Centroceras clavulatum, Ceramium flaccidum, Gelidium pusillum, Jania adhaerens, Jania ungulata e Polysiphonia scopulorum ocorreram em todas unidades amostrais do raso, enquanto que Galaxaura marginata e Hypnea musciformis ocorreram em todas as unidades amostrais do fundo.
Em termos de cobertura do estrato superior, foram identificadas duas faixas ao longo dos transetos: superior (0-3,0m), com domínio de calcárias articuladas (Corallinaceae); e inferior (3,1-5,0m), mais heterogênea, com abundância de Asparagopsis taxiformis e Sargassum spp., no entanto sem formar populações densas. Manchas de diferentes organismos mesclavam-se dentro das duas faixas. Crostas calcárias e Gelidiales sobressaíram na faixa de calcárias articuladas. As calcárias articuladas, Dictyota spp. e Laurencia spp. ocorreram ao longo dos transetos, em diferentes profundidades. Ouriços pretos (Echinometra lucunter) ocorreram na parte superior do costão (0,6-1,6m).
A área amostral deste ponto de coleta mostrou-se bastante heterogênea quanto aos aspectos fisionômicos, se comparada às demais.
Em termos de biomassa, as espécies mais representativas foram Sargassum filipendula (114,29g/m2- valor médio, no fundo), Hypnea musciformis (22,99g/m2- valor médio, no fundo) e Dictyota cervicornis (20,37g/m2- valor médio, no raso). As calcárias articuladas (Corallinaceae) foram mais expressivas no raso (valor médio= 287,47/m2), enquanto Galaxaura marginata e Tricleocarpa fragilis foram mais expressivas no fundo (valores médios= 171,41g/m2 e 3,84g/m2, respectivamente).
Coordenadas: 23º01’8,4”S e 44º25’53,7” W
Costão protegido do embate das ondas, de inclinação suave, sombreado durante parte do dia.
Profundidade: 5,2m
Temperatura à superfície: 29 ºC
Temperatura no fundo: 26 ºC
Salinidade: 37,5
Foram identificadas 47 espécies de algas bentônicas, sendo 10 Chlorophyta, 07 Phaeophyta e 30 Rhodophyta. Destas, as mais freqüentes foram Cladophora spp., Gelidium pusillum e Jania adhaerens, que ocorreram em unidades amostrais do raso e do fundo. Chondrachantus acicularis e Hypnea spinella ocorreram em todas as unidades amostrais do raso, enquanto Sargassum vulgare e S. filipendula e suas epífitas Dictyota cervicornis, Asparagopsis taxiformis- fase tetrasporofítica, Champia parvula, Hypnea musciformis e Pneophyllum fragile ocorreram em todas as unidades amostrais do fundo.
Em termos de cobertura do estrato superior, foram identificadas duas faixas ao longo dos transetos: superior (0-0,9m), com domínio de Gelidiales; e inferior (1,0-5,0m), com domínio de calcárias articuladas (Corallinaceae). Sobre o denso tapete de calcárias articuladas, algumas algas sobressaíram, como Dictyota spp., Ceramium spp. e Chondracanthus acicularis.
A área amostral deste ponto de coleta mostrou-se moderadamente heterogênea quanto ao aspecto fisionômico, se comparada às demais.
Em termos de biomassa, as espécies mais representativas foram Sargassum vulgare (24,53g/m2- valor médio, no fundo) e Dictyota cervicornis (9,97g/m2- valor médio, no fundo). As calcárias articuladas (Corallinaceae) foram mais expressivas no raso (valor médio= 236,90g/m2).
Coordenadas: 23º01’4,6”S e 44º26’9,1” W
Costão protegido do embate das ondas, de inclinação acentuada, sombreado durante parte do dia (manhã).
Profundidade: 6,0m
Foram identificadas 55 espécies de algas bentônicas, sendo 10 Chlorophyta, 07 Phaeophyta e 38 Rhodophyta. Destas, as mais freqüentes foram Amphiroa fragilissima, Ceramium flaccidum, Gelidiopsis planicaulis, Hypnea musciformis, Jania adhaerens, Polysiphonia subtilissima e Wrangelia argus, que ocorreram em unidades amostrais do raso e do fundo. Gelidium pusillum e Laurencia papillosa ocorreram em todas as unidades amostrais do raso, enquanto Sargassum vulgare e Polysiphonia denudata ocorreram em todas as unidades amostrais do fundo.
Em termos de cobertura do estrato superior, foram identificadas duas faixas ao longo dos transetos: superior (0-0,4m), com domínio de Gelidiales, especialmente Gelidium pusillum; e inferior (0,5-5,0m), com domínio de calcárias articuladas (Corallinaceae). Sobre o denso tapete de calcárias articuladas, algumas algas sobressaíram, como Ceramium spp., Chondracanthus acicularis e Hypnea spp.Plantas de Acanthophora spicifera apareceram isoladamente e em pequena quantidade na faixa de calcárias articuladas.
A área amostral deste ponto de coleta mostrou-se moderadamente heterogênea quanto ao aspecto fisionômico, se comparada às demais.
Em termos de biomassa, as espécies mais representativas foram Hypnea musciformis (12,37g/m2- valor médio, no raso), Gelidium pusillum (2,83g/m2- valor médio, no raso) e Wrangelia argus (1,23g/m2- valor médio, no fundo). As calcárias articuladas (Corallinaceae) foram mais expressivas no raso (valor médio= 16,11g/m2).
Coordenadas: 23º00’53,5”S e 44º26’32,1” W
Costão protegido do embate das ondas, com blocos rochosos de dimensões e inclinações variadas.
Profundidade: 4,7m
Temperatura à superfície: 32 ºC
Temperatura no fundo:26,5 ºC
Salinidade: 37
Foram identificadas 51 espécies de algas bentônicas, sendo 10 Chlorophyta, 09 Phaeophyta e 32 Rhodophyta. Destas, as mais freqüentes foram Dictyota cervicornis, Acanthophora spicifera, Amphiroa fragilissima, Asparagopsis taxiformis- fase tetrasporofítica, Hypnea spinella e Jania adhaerens, que ocorreram em unidades amostrais do raso e do fundo. Cladophora spp., Chondria spp. e Gelidium pusillum ocorreram em todas as unidades amostrais do raso, enquanto Sargassum vulgare, Ceramium flaccidum, Dasya spp., Galaxaura marginata, Gelidiopsis variabilis e Heterosiphonia crispella ocorreram em todas as unidades amostrais do fundo.
Em termos de cobertura do estrato superior, foram identificadas três faixas ao longo dos transetos: superior (0-1,3m), com domínio de Gelidiales; intermediária (1,4-2,5m), com domínio de calcárias articuladas (Corallinaceae); e inferior (2,6-5,0m), com domínio de Acanthophora spicifera. Na faixa de Gelidiales, sobressaíram também cracas e, sobre estas, algas filamentosas verdes (Cladophoraceae) e pardas (Ectocarpaceae). Sobre o denso tapete de calcárias articuladas, algumas algas sobressaíram, como Dictyota spp., Ceramium spp. e também Acanthophora spicifera. Mais abaixo, na faixa de Acanthophora spicifera, ocorreram principalmente Dictyota spp.e Padina spp. Plantas de Sargassum spp. foram observadas em blocos rochosos mais no fundo (fora da área abrangida pelos transetos), porém sem formar população densa.
A área amostral deste ponto de coleta mostrou-se moderadamente heterogênea quanto ao aspecto fisionômico, se comparada às demais.
Em termos de biomassa, as espécies mais representativas foram Sargassum vulgare (45,28g/m2- valor médio, no fundo), Acanthophora spicifera (20,85g/m2- valor médio, no raso) e Asparagopsis taxiformis- fase tetrasporofítica (14,40g/m2- valor médio, no fundo). As calcárias articuladas (Corallinaceae) foram mais expressivas no raso (valor médio= 213,17g/m2), enquanto Galaxaura marginata foi mais expressiva no fundo (valor médio= 62,99g/m2).
Costão protegido do embate das ondas, com blocos rochosos de pequenas dimensões e inclinação suave.
Profundidade: raso
Temperatura à superfície: 33 ºC
Temperatura no fundo: --
Salinidade: 35
Foram identificadas 22 espécies de algas bentônicas, sendo 10 Chlorophyta, 02 Phaeophyta e 10 Rhodophyta. Destas, as mais freqüentes foram Chaetomorpha brachygona, Cladophora spp., Sphacelaria furcigera, Sphacelaria tribuloides, Gelidium pusillum e Pterocladia bartlettii, que ocorreram nas três unidades amostrais. Muitas cianobactérias filamentosas (Oscillatoriaceae) foram coletadas, mas não identificadas em nível específico.
Em termos de cobertura do estrato superior, foi identificada apenas uma faixa, com domínio de filamentosas, especialmente cianobactérias e clorofíceas, que, em sua maioria, cresciam sobre cracas. Manchas de Gelidiales sobressaíram ao longo de toda a extensão dos transetos.
A área amostral deste ponto de coleta mostrou-se a menos heterogênea quanto ao aspecto fisionômico, se comparada às demais.
Em termos de biomassa, as espécies mais representativas foram Pterocladia bartlettii (54,77g/m2- valor médio, no raso) e Cladophora spp. (16,91g/m2- valor médio, no raso). Calcárias articuladas não foram representativas.
Costão moderadamente protegido ao embate das ondas, de inclinação acentuada.
Profundidade: --
Temperatura à superfície: 32o C
Temperatura no fundo: --
Salinidade: 35
Foram identificadas 46 espécies de algas bentônicas, sendo 08 Chlorophyta, 08 Phaeophyta e 30 Rhodophyta. Destas, as mais freqüentes foram Cladophora spp., Dictyota cervicornis, Acanthophora spicifera, Champia vieillardii, Gelidiopsis variabilis e Hypnea spinella, que ocorreram em unidades amostrais do raso e do fundo. Amphiroa fragilissima, Jania adhaerens e Laurencia papillosa ocorreram em todas as unidades amostrais do raso, enquanto Spatoglossum schroederi, Bryothamnion seaforthii, Galaxaura marginata, Gelidiopsis planicaulis, Heterosiphonia crispella e Hypnea musciformis ocorreram em todas as unidades amostrais do fundo. Cianobactérias filamentosas avermelhadas também foram muito freqüentes como epífitas em outras algas, porém não foram identificadas em nível específico.
Em termos de cobertura do estrato superior, foram identificadas três faixas ao longo dos transetos: superior (0-0,3m), com domínio de Laurencia papillosa e cracas; intermediária (0,4-3,4m), com domínio de Acanthophora spicifera e calcáriasarticuladas (Corallinaceae); e inferior (3,6-5,0m), com domínio da macrófita coriácea Bryothamnion seaforthii. Calcárias articuladas ocorreram em manchas também na faixa superior. Dictyota spp. e Gelidiales sobressaíram na faixa de Acanthophora spicifera. Na faixa de Bryothamnion seaforthii, sobressaíram Galaxaura marginata, Dictyopteris delicatula e Champia spp. Plantas de Sargassum, isoladas e muito epifitadas, foram observadas mais ao fundo (fora da extensão dos transetos).
A área amostral deste ponto de coleta mostrou-se moderadamente heterogênea quanto ao aspecto fisionômico, se comparada às demais.
Em termos de biomassa, as espécies mais representativas foram Bryothamnion seaforthii (134,99g/m2- valor médio, no fundo), Acanthophora spicifera (25,60g/m2- valor médio, no raso), Hypnea musciformis (11,31g/m2- valor médio, no fundo) e Dictyota cervicornis (6,29g/m2- valor médio, no raso). As calcárias articuladas (Corallinaceae) foram mais expressivas no raso (valor médio= 7,09g/m2), enquanto Galaxaura marginata foi mais expressiva no fundo (valor médio= 17,33g/m2).
Coordenadas: 23º00’16,6”S e 44º26’26,2” W
Costão moderadamente protegido ao embate das ondas, de inclinação moderada.
Profundidade: 6m
Temperatura à superfície: 31 ºC
Temperatura no fundo: 26 ºC
Salinidade: 36
Foram identificadas 47 espécies de algas bentônicas, sendo 07 Chlorophyta, 08 Phaeophyta e 32 Rhodophyta. Destas, as mais freqüentes foram Cladophora spp., Acanthophora spicifera, Amphiroa fragilissima, Asparagopsis taxiformis- fase tetrasporofítica, Ceramium flaccidum, Gelidium pusillum, Heterosiphonia crispella, Hypnea spinella, Jania adhaerens e Laurencia papillosa, que ocorreram em unidades amostrais do raso e do fundo.Cianobactérias filamentosas avermelhadas (Oscillatoriaceae) também foram muito freqüentes como epífitas em outras algas, porém não foram identificadas em nível específico. Dictyota cervicornis, Sargassum filipendula, Champia vieillardii, Chondracanthus acicularis, Dasya spp., Galaxaura marginata, Gelidiopsis planicaulis e Polysiphonia ferulacea ocorreram em todas as unidades amostrais do fundo.
Em termos de cobertura do estrato superior, foram identificadas três faixas ao longo dos transetos: superior (0-0,3m), com domínio de Laurencia papillosa; intermediária (0,4- 4,4m), com domínio de calcárias articuladas (Corallinaceae) e Acanthophora spicifera; e inferior (4,5-5,0m), com domínio de Galaxaura marginata. Na faixa de Laurencia papillosa, foi registrada a ocorrência expressiva de filamentosas verdes. Na faixa de calcárias articuladas e Acanthophora spicifera, sobressaíram Gelidium pusillum (0,4-4,1m do transeto), filamentosas vermelhas (Ceramiaceae) (1,5-4,4m do transeto), Dictyota cervicornis (2,4-4,1m do transeto) e Padina gymnospora (3,1-3,7m do transeto). Na faixa de Galaxaura marginata, ocorreram também calcárias articuladas e Dictyota spp; plantas de Sargassum spp. cresciam isoladamente ou em pequenas manchas.
A área amostral deste ponto de coleta mostrou-se moderadamente heterogênea quanto ao aspecto fisionômico, se comparada às demais.
Em termos de biomassa, as espécies mais representativas foram Sargassum filipendula (29,39g/m2- valor médio, no fundo), Sargassum vulgare (12,80g/m2- valor médio, no fundo), Hypnea spinella (5,81g/m2- valor médio, no fundo) e Gelidiopsis variabilis (5,07g/m2- valor médio, no fundo). As calcárias articuladas (Corallinaceae) foram mais expressivas no raso (valor médio= 377,76g/m2), enquanto Galaxaura marginata foi mais expressiva no fundo (valor médio= 112,53g/m2).
- Área sob influência direta do efluente - Saco Piraquara de Fora - Ponta de Pingo d’Água (Ponto B.52 do “pré-operacional”)
Coordenadas: 23º00’6,8”S e 44º26’7,6” W
Costão moderadamente exposto ao embate das ondas, de inclinação moderada.
Profundidade: 6m
Temperatura à superfície: 30o C
Temperatura no fundo: 26o C
Salinidade: 36
Foram identificadas 60 espécies de algas bentônicas, sendo 13 Chlorophyta, 09 Phaeophyta e 38 Rhodophyta. Destas, as mais freqüentes foram Cladophora spp., Acanthophora spicifera, Gelidiopsis variabilis, Hypnea spinella e Jania adhaerens, que ocorreram em unidades amostrais do raso e do fundo.Cianobactérias filamentosas (Oscillatoriaceae) foram freqüentes, mas não foram identificadas em nível específico. Amphiroa fragilissima e Gelidium pusillum ocorreram em todas as unidades amostrais do raso, enquanto Sargassum vulgare, Asparagopsis taxiformis-fase tetrasporofítica, Ceramium flaccidum, Champia parvula, Champia vieillardii, Hypnea musciformis e Pneophyllum fragile ocorreram em todas as unidades amostrais do fundo.
Em termos de cobertura do estrato superior, foram identificadas duas faixas ao longo dos transetos: superior (0-1,6m), com domínio de Gelidiales; inferior (1,7-5,0m), com domínio de calcárias articuladas (Corallinaceae). Calcárias articuladas também foram abundantes na faixa superior, juntamente com filamentosas verdes (Cladophoraceae) e vermelhas (Ceramiaceae) e Dictyota spp. Na faixa inferior, no meio do banco das calcárias articuladas, cresciam plantas de Padina gymnospora, Gelidiales e Acanthophora spicifera, que não formavam populações densas.
A área amostral deste ponto de coleta mostrou-se pouco heterogênea quanto ao aspecto fisionômico, se comparada às demais.
Em termos de biomassa, as espécies mais representativas foram Sargassum vulgare (80,96g/m2- valor médio, no fundo), Sargassum filipendula (21,71g/m2- valor médio, no fundo), Champia vieillardii (9,28g/m2- valor médio, no fundo) e Hypnea musciformis (8,27g/m2- valor médio, no fundo). As calcárias articuladas (Corallinaceae) foram mais expressivas no raso (valor médio= 179,52g/m2), enquanto Galaxaura marginata foi mais expressiva no fundo (valor médio= 14,72g/m2).
- Área sob influência direta do efluente - Saco Piraquara de Fora - Ilha de Pingo d’Água, lado voltado para o continente, atrás do “píer”
Coordenadas: 23º00’6,0”S e 44º25’52,3” W
Costão protegido do embate das ondas, de inclinação acentuada.
Profundidade: 7,0m
Temperatura à superfície: 29,5o C
Temperatura no fundo: 26o C
Salinidade: 36,5
Foram identificadas 45 espécies de algas bentônicas, sendo 05 Chlorophyta, 08 Phaeophyta e 32 Rhodophyta. Destas, as mais freqüentes foram Dictyota cervicornis, Amphiroa fragilissima, Ceramium flaccidum e Hypnea spinella, que ocorreram em unidades amostrais do raso e do fundo.Cianobactérias filamentosas (Oscillatoriaceae), não identificadas em nível específico, foram freqüentes no raso. Pseudoderbesia arbuscula, Jania adhaerens e Laurencia papillosa ocorreram em todas as unidades amostrais do raso, enquanto Padina gymnospora, Dasya spp. e Galaxaura marginata ocorreram em todas as unidades amostrais do fundo.
Em termos de cobertura do estrato superior, foram identificadas duas faixas ao longo dos transetos: superior (0-0,7m), com domínio de Laurencia papillosa; inferior (0,8-5,0m), com domínio de calcárias articuladas (Corallinaceae). Calcárias articuladas também ocorreram na faixa superior, junto com Ulva spp., Gelidium pusillum e Ceramiaceae. Na faixa inferior, sobressaíram Dictyota spp., Padina gymnospora, Laurencia sp. e Ceramiaceae. Acanthophora spicifera, embora presente, não formava populações densas. Plantas de Sargassum spp. foram observadas mais ao fundo (fora da extensão dos transetos).
A área amostral deste ponto de coleta mostrou-se moderadamente heterogênea quanto ao aspecto fisionômico, se comparada às demais.
Em termos de biomassa, as espécies mais representativas foram Dictyota cervicornis (16,69g/m2 e 5,44g/m2 - valores médios, no raso e no fundo, respectivamente), Padina gymnospora (2,61g/m2- valor médio, no fundo). As calcárias articuladas (Corallinaceae) foram mais expressivas no raso (valor médio= 157,92g/m2), enquanto Galaxaura marginata foi mais expressiva no fundo (valor médio= 17,07g/m2).
- Área controle - Saco Piraquara de Dentro - Ponta do Pasto
Costão moderadamente exposto ao embate das ondas, de inclinação moderada.
Foram identificadas 65 espécies de algas bentônicas, sendo 09 Chlorophyta, 12 Phaeophyta e 44 Rhodophyta. Destas, as mais freqüentes foram Sargassum vulgare, Acanthophora spicifera, Amphiroa fragilissima, Chondracanthus acicularis, Hypnea musciformis e Jania adhaerens, que ocorreram em unidades amostrais do raso e do fundo.Cianobactérias filamentosas (Oscillatoriaceae), não identificadas em nível específico, também foram freqüentes. Ceramium brasiliense ocorreu em todas as unidades amostrais do raso, enquanto Dictyopteris delicatula, Dictyota cervicornis, Sargassum filipendula, Champia parvula e Dasya spp. ocorreram em todas as unidades amostrais do fundo.
Em termos de cobertura do estrato superior, foram identificadas três faixas ao longo dos transetos: superior (0-0,2m), com domínio de filamentosas pardas (Ectocarpaceae) e vermelhas (Ceramiaceae) e cracas; intermediária (0,3-3,8m), com domínio de calcárias articuladas (Corallinaceae); inferior (3,9-5,0m), com domínio de Sargassum spp. A faixa intermediária mostrou-se como a mais heterogênea, onde diferentes algas cresciam dentre as almofadas das calcárias articuladas, isoladamente ou em pequenas manchas, especialmente Dictyota spp., Padina gymnospora, Hypnea spp., Laurencia spp., Gelidiales e Ceramiaceae. Calcárias articuladas, no entanto, ocorreram também nas demais faixas. Na faixa inferior, as plantas de Sargassum não formavam população densa, sendo epifitadas por diferentes macroalgas. Nesta faixa, sobressaíram Laurencia spp., algas crostosas rosas e animais sésseis, como esponjas e cracas.
A área amostral deste ponto de coleta mostrou-se muito heterogênea quanto ao aspecto fisionômico, se comparada às demais.
Em termos de biomassa, as espécies mais representativas foram Sargassum filipendula (68,21g/m2 - valor médio, no fundo), Sargassum vulgare (10,99g/m2 - valor médio, no fundo), Dictyopteris delicatula (3,20g/m2- valor médio, no fundo) e Dictyota cervicornis (3,04g/m2- valor médio, no fundo). As calcárias articuladas (Corallinaceae) foram mais expressivas no raso (valor médio= 60,64g/m2), enquanto Galaxaura marginata foi mais expressiva no fundo (valor médio= 9,33g/m2).
Costão protegido do embate das ondas, de inclinação moderada.
Foram identificadas 71 espécies de algas bentônicas, sendo 14 Chlorophyta, 10 Phaeophyta e 47 Rhodophyta. Destas, as mais freqüentes foram Cladophora spp., Dictyota cervicornis, Acanthophora spicifera, Asparagopsis taxiformis - fase tetrasporofítica, Gelidiopsis variabilis, Hypnea spinella, Jania adhaerens, Pneophyllum fragile e Wrangelia argus, que ocorreram em unidades amostrais do raso e do fundo. Cianobactérias filamentosas (Oscillatoriaceae), não identificadas em nível específico, foram freqüentes no raso. Ceramium flaccidum, Dasya spp. e Laurencia papillosa ocorreram em todas as unidades amostrais do raso, enquanto Sargassum vulgare, Champia parvula, Champia vieillardii, Polysiphonia denudata e P. ferulacea ocorreram em todas as unidades amostrais do fundo.
Em termos de cobertura do estrato superior, foram identificadas três faixas ao longo dos transetos: superior (0-0,3m), com domínio de Gelidium pusillum; intermediária (0,4-2,8m), com domínio de Acanthophora spicifera; inferior (2,9-5,0m), com domínio de Sargassum vulgare. Na faixa superior, filamentosas pardas (Ectocarpaceae) também foram importantes em termos de cobertura. A faixa intermediária mostrou-se como a mais heterogênea, onde diferentes algas cresciam junto com Acanthophora spicifera, principalmente Dictyota spp., Padina spp., Sargassum vulgare, calcárias articuladas e Ceramiaceae. Calcárias articuladas, no entanto, não foram dominantes em nenhuma faixa. Na faixa inferior, as plantas de Sargassum formavam população densa, com plantas muito desenvolvidas; sob Sargassum ou como epífitas, sobressaíram Dictyota spp., Padina spp., Ceramiaceae e Acanthophora spicifera.
A área amostral deste ponto de coleta mostrou-se moderadamente heterogênea quanto ao aspecto fisionômico, se comparada às demais.
Em termos de biomassa, as espécies mais representativas foram Sargassum vulgare (210,40g/m2 - valor médio, no fundo), Acanthophora spicifera (88,59g/m2 - valor médio, no raso) e Laurencia papillosa (7,63g/m2- valor médio, no raso). As calcárias articuladas (Corallinaceae) foram mais expressivas no raso (valor médio= 4,53g/m2).
Biomassa dos Táxons mais Representativos
Na área da descarga do efluente (B 32), só foram representativas, em termos de biomassa, espécies de Cladophora, Sphacelaria (filamentosas) e Gelidiales (macrófita corticada). Nas áreas de influência direta dos efluentes (Saco Piraquara de Fora) e controle, outras espécies de macroalgas, pertencentes a diferentes tipos morfo-funcionais, mostraram biomassa expressiva. Calcárias articuladas foram bem representadas em pontos de coleta desta área: Corallinaceae, principalmente no nível raso dos pontos de coleta do Saco Piraquara de Fora; espécies de Galaxaura, no nível fundo. Representantes de macrófitas corticadas e coriáceas foram bastante variáveis, dependendo do ponto de coleta, particularmente no caso de Acanthophora spicifera e Sargassum spp., que ora ocorreram com biomassa expressiva, ora não.
A maioria das espécies representativas, em termos de biomassa, alcançou seus máximos valores médios, por nível, em pontos de coleta da área controle; exceção para as Corallinaceae e Hypnea spinella.
A seguir são relacionados os táxons de macroalgas que ocorreram com biomassa representativa na maioria dos pontos de coleta da área de estudo e onde alcançaram seus valores máximos, em média (Tabela 52).
Tabela 52 - Táxons com ocorrência representativa na maioria dos pontos da Área de Estudo
Os maiores valores de biomassa de Sargassum spp. no nível fundo podem ser relacionados ao fato de ser local mais estável, não sujeito ao estresse provocado pelos períodos de emersão de níveis superiores no costão, o que está de acordo com o observado por outros autores (Umezaki, 1984; Hurtado & Ragaza, 1999).
Mesmo os maiores valores de biomassa de Sargassum, observados em pontos de coleta da área controle, mostraram-se inferiores aos mencionados para outras espécies de regiões tropicais e sub-tropicais: Sargassum spp. para o sudeste do Brasil (491,00g/m2) (Oliveira Filho & Paula, 1979); S. cymosum var. cymosum de Ubatuba, São Paulo (800g/m2) (Paula & Oliveira Filho, 1980); S. cymosum var. cymosum de Bombinhas, Santa Catarina (411,88g/m2) (Dubiaski-Silva & Masunari, 2000); Sargassum spp. de Curaçao (440,00g/m2) (Wanders, 1976); Sargassum sppdas Filipinas (3275,00g/m2) (Cordero Jr., 1981); Sargassum spp. de Magnetic Island, Austrália (700g/m2) (Vuki & Price, 1994).
Comparando-se a biomassa das espécies mais representativas da Ponta da Fortaleza, Ponta de Pingo d’Água e Ponta da Pitanga, com os dados fornecidos por Széchy (1996), para bancos de Sargassum dos mesmos pontos de coleta, estudados anteriormente, podemos constatar claramente a diminuição da biomassa alcançada pelas populações de Sargassum dos três pontos de coleta, principalmente na Ponta de Pingo d’Água, onde simultaneamente houve um aumento considerável na biomassa das calcárias articuladas.
Para as outras espécies, não é possível traçar uma tendência para esta variável. Nesse aspecto, há de ser salientado que é realmente esperada a variação de biomassa, entre pontos de coleta e mesmo entre épocas do ano, nas espécies de tipo morfo-funcional mais simples, que não representam as dominantes na comunidade, tendo, muitas vezes, comportamento oportunista. A Tabela 53 compara a biomassa (peso seco, g/m2) das espécies de macroalgas mais representativas, em pontos de coleta da Baía da Ribeira, em 1991-1992 e em 2002.
Tabela 53 - Comparação da biomassa (peso seco, g/m2) das espécies de macroalgas mais representativas, na Área de Estudo
Clique aqui para visualizar a tabela 53
Para fins de monitoramento, é importante avaliar a variação de biomassa de espécies perenes e dominantes nos costões rochosos. No caso da área de estudo, esta variável deve ser acompanhada para os representantes de Sargassum e Acanthophora, como já havia sido recomendado no “pré-operacional”, como também das Corallinaceae, Gelidiales e cianobactérias, dentre outras.
Ênfase deve ser dada aos estudos de Sargassum, por sua indubitável importância ecológica e por sua ampla distribuição na Baía da Ilha Grande e em vários outros pontos do litoral do Brasil, o que possibilita comparações do comportamento das espécies em diferentes condições ambientais.
Sargassum é elemento conspícuo, em função de sua ocorrência e abundância, em comunidades marinhas bentônicas de várias localidades do Planeta, especialmente em regiões tropicais e sub-tropicais (De Wreede, 1976; Shunula, 1988; Eston & Bussab, 1990; Vuki & Price, 1994; Stiger & Payri, 1999). Mesmo em regiões tropicais e sub-tropicais, onde não se reconhecem com nitidez as estações do ano, são descritas variações temporais nas populações de Sargassum (Paula & Oliveira Filho, 1980; Cordero Jr., 1981; Shunula, 1985; De Ruyter van Steveninck & Breeman, 1987; Trono Jr. & Lluisma 1990; Largo et al., 1994; Hurtado & Ragaza, 1999). Tais variações são verificadas principalmente na altura e biomasa das plantas, na quantidade de ramos estéreis e férteis, na quantidade de folhas e de receptáculos. Para a maioria das espécies de Sargassum, que é perene, podem-se definir diferentes fases na vida das plantas adultas, a saber: crescimento vegetativo, pico reprodutivo, senescência, queda de ramos e folhas e regeneração (Ang Jr., 1985a). A regeneração é um processo muito importante para a manutenção das plantas na comunidade, ocorrendo através do apressório e dos ramos principais (Ang Jr., 1985b).
Para a área de estudo, variações temporais de abundância nas populações de Sargassum já haviam sido referidas. No período “pré-operacional”, a biomassa das populações de Sargassum foi mínima em julho e setembro de 1980, e máxima, em abril de 1980 (FUJB, 1980/81). Após o início de funcionamento da Unidade I, foi descrito também o declínio nas populações nos meses de inverno (Széchy, 1996).
Na literatura, o declínio da abundância de Sargassum é relacionado a diferentes fatores, principalmente às variações de temperatura da água do mar (Ang Jr., 1986; Shunula, 1988; Vuki & Price, 1984; Hurtado & Ragaza, 1999). Além da temperatura, são citados: fotoperíodo, concentração de nutrientes, salinidade, grau de exposição às ondas (Széchy & Paula, 2000b), nível das marés (Paula & Oliveira Filho, 1980), ressacas (De Ruyter van Steveninck & Breeman, 1987), epifitismo (Dubiaski-Silva & Masunari, 2000) e herbivoria. Contudo, não há um consenso sobre os processos e como estes fatores controlam seu ciclo de vida.
Cordero Jr. (1981) conclui que temperaturas acima de 28-30ºC favorecem o crescimento de espécies de Sargassum spp das Filipinas. Por outro lado, McCourt (1984) relaciona o declínio na abundância de espécies de Sargassum do norte do Golfo da Califórnia no período de 1977-1981, às temperaturas quentes da água do mar no ano de 1978. Para Glenn et al. (1990), variações significativas de biomassa em Sargassum podem ser explicadas por pequenas variações sazonais de temperatura da água do mar (~=5 ºC), como o observado em espécies do Havaí. De um modo geral, quanto à sazonalidade, há dois grupos de espécies perenes de Sargassum: aquelas com crescimento vegetativo máximo no inverno (McCourt, 1984; Espinoza & Rodríguez, 1989) e aquelas com crescimento máximo no verão (Ang Jr., 1985).
Com base no conhecimento de aspectos da biologia de diferentes espécies de Sargassum, fornecido pela literatura e sintetizados acima, podemos concluir que o aumento da temperatura da água do mar na área de estudo é um fator de importância primária para explicar a variação de biomassa das populações de Sargassum.Sobretudo, deve-se considerar a influência da temperatura da água do mar sobre o comportamento de outros fatores abióticos e bióticos, e a ação sinérgica dos mesmos.
A Figura 62 fornece os valores médios e os desvios-padrão da biomassa (peso seco, em g/m2) das macroalgas mais representativas na área de estudo, agrupadas por tipo morfo-funcional, coletadas nos níveis raso e fundo dos 12 pontos de coleta.
Macrófitas corticadas, especialmente Acanthophora spicifera, e macrófitas coriáceas, especialmente Sargassum spp., apresentaram os maiores valores de biomassa na área controle (em locais protegidos), enquanto as filamentosas apresentaram os maiores valores de biomassa no local da descarga (Ponto B.32). As calcárias articuladas mostraram os maiores valores de biomassa em alguns pontos de coleta da área de influência direta do efluente (Saco Piraquara de Fora).
De um modo geral, as calcárias articuladas e as macrófitas corticadas foram mais abundantes no nível raso dos costões rochosos, enquanto as macrófitas coriáceas foram mais abundantes no nível fundo. Filamentosas e foliáceas não mostraram um comportamento uniforme quanto a este aspecto, sendo mais abundantes nos níveis raso ou fundo, de acordo com o ponto de coleta.
Segundo Steneck & Dethier (1994), a abundância de algas, avaliada em termos de biomassa, varia em função do grau de distúrbios, medido pela sua freqüência e intensidade. Grupos morfo-funcionais mais complexos, especialmente o das macrófitas coriáceas, como Sargassum spp., são mais abundantes em locais com baixo grau de distúrbios. À medida que o grau de distúrbios aumenta, a biomassa das macrófitas coriáceas diminui até que estas desaparecem e são substituídas por algas foliáceas e filamentosas.
Em situações extremas, aparecem apenas representantes de microalgas e de algas crostosas, com biomassa significativamente menor (Steneck & Dethier, 1994). Lambert & Steinke (1986) e Dye (1992) mostram a relação entre a ocorrência de distúrbios e o domínio de calcárias articuladas nas comunidades bentônicas de costões rochosos. Eston & Bussab (1990) e Fonseca (1998) também relacionam distúrbios ao aparecimento de outros tipos morfo-funcionais de macroalgas, no caso foliáceas, em comunidades bentônicas antes dominadas por macrófitas coriáceas.
Com base nas descrições de padrões estruturais quantitativos para bancos de Sargassum do litoral dos estados do Rio de Janeiro e São Paulo (Széchy, 1996; Széchy & Paula, 2000b), levanta-se a hipótese de que as mudanças provocadas no ambiente marinho da área sob influência direta do efluente da CNAAA devem estar afetando o processo de colonização e sucessão nessas comunidades, que parecem não estar tendo condições para atingir o clímax, caracterizado pelo domínio de macrófitas corticadas, especificamente de Sargassum. O domínio de calcárias articuladas em muitas dessas comunidades pode ser interpretado como um estadio intermediário destas comunidades. Deve-se estudar com cautela até que ponto esta situação é reversível ou permanente.
Figura 62 - Biomassa (peso seco, g/m2) das macroalgas agrupadas por tipo morfo-funcional, nos 12 pontos de coleta, por nível do costão. Valores médios e desvios-padrão.
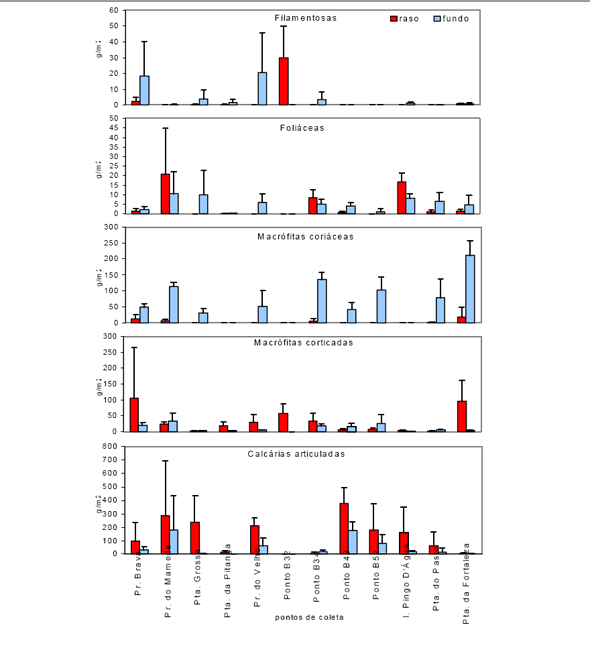
Influência da temperatura da água do mar
Os resultados apresentados (padrões de zonação, composição e riqueza específicas de macroalgas nos pontos de coleta e nos diferentes níveis do costão, freqüência e biomassa das espécies) nos levam a indicar a influência da temperatura da água do mar como um dos fatores abióticos mais importantes, nas mudanças detectadas nas comunidades fitobentônicas, quando considerados os dados pretéritos. Evidentemente, não se pode deixar de mencionar o papel conjunto de vários fatores na estruturação das comunidades bentônicas, incluindo outros fatores abióticos e também fatores bióticos. Estes fatores interagem de modo complexo, sendo, portanto, difícil delimitar até onde vão os efeitos de cada um isoladamente.
A título de exemplificação de outro fator abiótico de provável influência na comunidade fitobentônica da área de estudo, destaca-se a presença de compostos químicos adicionados ao efluente, conforme o apontado para comunidades do Mar Báltico por Lehtinen et al. (1988). Com base em experimentos no laboratório e no mar, esses autores mostraram que a alga parda Fucus vesiculosus L. (Fucales) é extremamente sensível à exposição prolongada ao cloro dos efluentes das fábricas de papel. Embora o efeito do cloro sobre a alga seja restrito no tempo e no espaço, seus efeitos secundários podem se estender consideravelmente, ao diminuir a taxa de recrutamento em populações de outros organismos vinculados ao Fucus vesiculosus, através da cadeia alimentar. Sendo Sargassum (Fucales), a alga dominante na região sublitorânea rasa de muitos costões rochosos da área de estudo, bem como do litoral sudeste do Brasil (Eston & Bussab, 1990; Széchy & Paula, 2000), há de se destacar a importância da avaliação dos efeitos de compostos químicos dissolvidos nos efluentes das unidades 1 e 2 sobre seu crescimento e reprodução.
Com base em dados da literatura, podemos relacionar aspectos qualitativos e quantitativos descritos para as comunidades bentônicas da área de estudo ao aumento da temperatura da água do mar, constatado especialmente na massa d’água superficial do Saco Piraquara de Fora.
É indiscutível que a distribuição geográfica atual das macroalgas no ambiente marinho é regulada pelas históricas variações da temperatura da água do mar, dentre outros fatores (Lüning, 1990). A temperatura da água do mar pode ser um fator de estresse fisiológico para as macroalgas marinhas (Lobban & Harrison, 1995), que a ele respondem de forma reversível, se removido o estresse, ou de forma permanente, resultando em injúrias, mesmo quando interrompido o estresse (Levitt, 1972).
É difícil determinar com precisão o nível de estresse em que se iniciam as injúrias nas macroalgas; também é difícil reconhecer o exato nível de estresse que pode acarretar sua morte. Para a determinação destes níveis, são necessários estudos experimentais de tolerância para cada espécie de macroalga ou mesmo para cada linhagem. Infelizmente, os estudos existentes concentram-se nos efeitos de temperaturas muito baixas, em espécies de macroalgas de regiões do Hemisfério Norte (Cambridge et al., 1984; Lüning, 1984; Yarish et al., 1984; Lüning & Freshwater 1988). Em relação aos efeitos de temperaturas elevadas em macroalgas, ressaltamos os estudos realizados para algumas algas pardas, como para espécies de Fucales de regiões entre-marés (Schonbeck & Norton, 1978) e de Laminariales da região sublitorânea (Andrews, 1976), que concluíram que a capacidade de regeneração e de sobrevivência das espécies diminuiu com o tempo de exposição.
Em síntese, podem ser apontados os seguintes efeitos de temperaturas elevadas sobre as macroalgas marinhas: (i) alterações no crescimento das plantas; e (ii) alterações na reprodução das plantas.
A taxa de crescimento das plantas pode ser afetada por diferentes mecanismos, induzidos pelo aumento de temperatura: desnaturação de proteínas, destruição de enzimas termolábeis, destruição de membranas, com efeito direto no fotossistema II da fotossíntese (PSII) e na respiração (Soeder & Stengel, 1974). Altas temperaturas também podem levar ao aparecimento de doenças microbianas, que afetam o desenvolvimento das plantas (Andrews, 1976).
Segundo Lüning (1990), para espécies de macroalgas marinhas de regiões temperadas quentes e tropical, a temperatura ótima para o crescimento vegetativo está entre 15-30oC; além disso, espécies destas regiões, que habitam a região sublitorânea, parecem ter os mesmos limites de sobrevivência a temperaturas elevadas: 33-35oC. No entanto, Johannes & Betzer (1975) afirmam que o lançamento de água aquecida pode produzir mais estresse nos trópicos do que em regiões temperadas. Segundo esses autores, nos trópicos, comunidades marinhas da região sublitorânera vivem em ambiente relativamente estável, onde as variações sazonais de temperatura são, em média, menores. Com isso, acreditam que as espécies não tenham desenvolvido significante resistência a flutuações nas características físicas do ambiente. Talvez seja essa uma razão para sua baixa tolerância a poluentes, de um modo geral.
A temperatura da água do mar, juntamente com o fotoperíodo, influencia o tipo de reprodução que uma alga irá realizar, ou seja, determina o tipo de elemento de reprodução que irá ser formado: esporos ou gametas (Dring, 1974). Existem muitos estudos, com base em cultivo em laboratório, sobre o efeito destes fatores no ciclo de vida de espécies de macroalgas marinhas, particularmente de feofíceas (Müller, 1962; Kornmann, 1962). Entretanto, o controle do processo reprodutivo nas macroalgas, no ambiente onde vivem, está longe de ser integralmente compreendido pelo Homem. Aos efeitos da temperatura e da luz sobre o tipo de reprodução, acrescentam-se outros fatores, inclusive o controle endógeno, ou seja, os chamados ritmos circadianos (Dring, 1974).
O efeito da temperatura na quantidade de elementos de reprodução formados foi estudado para algumas espécies de macroalgas, mas de modo não conclusivo (Christie & Evans, 1962; Bellis, 1968; Hsiao, 1970; Sears & Wilce, 1970).
Há lacunas no conhecimento sobre os processos de crescimento e reprodução das macroalgas marinhas, bem como sobre os processos que regulam a estrutura e dinâmica das comunidades fitobentônicas, especialmente as dos trópicos. Mas esta constatação não impede de se afirmar que alterações nas taxas de crescimento e de reprodução das espécies de uma comunidade bentônica podem modificar, de modo diferencial, sua capacidade de dispersão, colonização e competição. Nesse contexto, algumas espécies podem ser eliminadas da comunidade, enquanto outras podem adaptar-se às novas condições, tornando-se dominantes. Isto significa que são esperadas alterações, a médio e longo prazos, na estrutura de comunidades fitobentônicas, submetidas ao estresse de aumento de temperatura.
A adaptação ao estresse por temperatura pode se dar por diferentes estratégias, dependendo de características das espécies (estenotérmicas x euritérmicas; tipos morfo-funcionais diferentes; ciclos de vida diferentes) e do ambiente (região entre-marés x região sublitorânea; região tropical x região temperada).
Dentre as estratégias de adaptação das macroalgas ao estresse por temperatura, podemos citar, como exemplos: a alternância de gerações heteromórficas frente às variações espaciais e temporais na temperatura da água do mar; o ciclo de vida mais rápido de formas morfologicamente mais simples, como filamentosas; a reprodução exclusivamente assexuada por propágulos, em locais com temperaturas mais elevadas. Lüning (1990) cita também a possibilidade de adaptação fisiológica das espécies a temperaturas mais elevadas através do processo gradual de diferenciação ecotípica ou através de mutações. Parece que algumas espécies criam resistência ao estresse térmico através da síntese de proteínas especiais (Lai et al., 1988).
Considerando o indiscutível papel das macroalgas na cadeia alimentar de regiões costeiras, além de seu papel como refúgio e local de reprodução para diferentes espécies da fauna marinha, é oportuno e urgente questionarmos em que intensidade e como as alterações na estrutura das comunidades fitobentônicas serão repassadas para outros componentes do ecossistema. Portanto, são necessários ainda muitos estudos, de diferentes espécies de macroalgas e com diferentes abordagens.
7.2.3.2. Análise das Regiões Estudadas - topo
A flora da área de estudo, baseada no levantamento dos dados pretéritos (251 espécies) e no atual levantamento, pode ser considerada rica.
Cerca de 73% das espécies mencionadas no estudo pré-operacional, foram novamente coletadas, aparentemente, indicando que não houve a exclusão ou a substituição de parcela significativa das espécies da flora marinha da área.
Quando analisados individualmente, os 12 pontos de coleta apresentam uma marcante variação no número de espécies.
Existe uma tendência à diminuição do número de espécies da área controle em direção até o ponto de descarga dos efluentes (B. 32) e simultaneamente, a diminuição do número de espécies no nível “raso” do costão (temperatura mais elevada), na área sob influência direta destes efluentes (Saco Piraquara de Fora).
O número de espécies de Phaeophyta, grupo considerado bom indicador da qualidade do ambiente marinho, diminuiu drasticamente do Saco Piraquara de Dentro (área controle) para o local da descarga, no Saco Piraquara de Fora (Ponto B.32).
Em relação ao nível do costão, o maior número de espécies foi encontrado no fundo (Ponto B.43, com 40 espécies), enquanto o menor foi encontrado no raso (Ponto B.32, com 8 espécies).
A maioria das espécies representativas, em termos de biomassa, alcançou seus máximos valores médios, por nível, em pontos de coleta da área controle; exceção para as calcárias articuladas e Hypnea spinella.
Os resultados apresentados (padrões de zonação, composição e riqueza específicas de macroalgas nos pontos de coleta e nos diferentes níveis do costão, freqüência e biomassa das espécies) nos levam a indicar a influência da temperatura da água do mar como um dos fatores abióticos mais importantes, nas mudanças detectadas nas comunidades fitobentônicas, quando considerados os dados anteriores.
7.3. Fauna Terrestre - topo
7.3.1. Herpetofauna - topo
No que se refere à herpetofauna da região da costa verde, nenhum grande estudo foi publicado até hoje, embora alguns autores tenham descrito ou citado espécies para a região. No Plano de Manejo da Estação Ecológica de Tamoios (Fase 1) é citada a ocorrência de espécies de 3 gêneros que não foram identificados, assim como a inexistência de trabalhos na área.
A seguir é apresentada uma lista de espécies encontradas na literatura ou em algumas coleções zoológicas (Tabela 54). As principais coleções zoológicas consultadas foram: ZUFRJ - Coleção de Anfíbios e Répteis do Instituto de Biologia da Universidade Federal do Rio de Janeiro; EI - Coleção Herpetológica Eugênio Izecksohn depositada no Instituto de Biologia da Universidade Federal Rural do Rio de Janeiro; e UNIRIO - Coleção de Anfíbios e Répteis da Escola de Ciências Biológicas da Universidade do Rio de Janeiro.
Tabela 54 - Lista de Anfíbios para a Região de Angra dos Reis e Proximidades
| Espécie |
Familia |
| Dendrophryniscus brevipollicatus |
BUFONIDAE |
| Brachycephalus vertebralis |
BRACHYCEPHALIDAE |
| Brachycephalus ephippium |
BRACHYCEPHALIDAE |
| Psyllophryne didactyla |
BRACHYCEPHALIDAE |
| Psyllophryne hermogenesii |
BRACHYCEPHALIDAE |
| Hyla arildae |
HYLIDAE |
| Hyla callypigia |
HYLIDAE |
| Hyla clepsydra |
HYLIDAE |
| Scinax angrensis |
HYLIDAE |
| Scinax ariadne |
HYLIDAE |
| Scinax argyreornatus |
HYLIDAE |
| Scinax atratus |
HYLIDAE |
| Scinax humilis |
HYLIDAE |
| Scinax perpusillus |
HYLIDAE |
| Scinax trapicheiroi |
HYLIDAE |
| Sphaenorhynchus orophilus |
HYLIDAE |
| Cyclorhamphus boraceiensis |
LEPTODACTYLIDAE |
| Cyclorhamphus eleutherodactylus |
LEPTODACTYLIDAE |
| Cyclorhamphus granulosus |
LEPTODACTYLIDAE |
| Hylodes phyllodes |
LEPTODACTYLIDAE |
| Megaelosia bocainensis |
LEPTODACTYLIDAE |
| Paratelmatobius gaigeae |
LEPTODACTYLIDAE |
| Paratelmatobius pictiventris |
LEPTODACTYLIDAE |
| Physalaemus barrioi |
LEPTODACTYLIDAE |
| Proceratophrys appendiculata |
LEPTODACTYLIDAE |
| Eleutherodactylus bolbodactylus |
LEPTODACTYLIDAE |
| Eleutherodactylus parvus |
LEPTODACTYLIDAE |
| Eleutherodactylus pusillus |
LEPTODACTYLIDAE |
| Thoropa miliaris |
LEPTODACTYLIDAE |
| Zachaenus parvulus |
LEPTODACTYLIDAE |
7.3.1.1. Caracterização da Herpetofauna - topo
Com atividades diurnas e noturnas, foram realizadas incursões para análise das condições ambientais e aspectos dos habitats preferenciais das espécies de anfíbios e répteis na área de influência da Usina Angra 3. Estabeleceram-se cronologicamente nove pontos de coleta e observação (Figura 63). Visitaram-se três ilhas da região próximas ao Saco Piraquara de Fora: Ilha do Pingo d'Água, Ilha do Brandão e Ilha de Paquetá. Nestas foram feitas observações apenas diurnas.
Nas ilhas, foram analisadas principalmente a formação rochosa, a cobertura vegetal, o folhiço da mata (serapilheira), o sistema de drenagem, os córregos e/ou acúmulos de água com possibilidade de vida para determinadas espécies de anfíbios e répteis.
Tanto na costa quanto nas ilhas, foram feitas entrevistas com os moradores locais sobre a ocorrência de repteis da região. A captura de repteis foi feita manualmente ou com auxílio de ganchos, principalmente para coleta de serpentes. Para procura de anfíbios de chão de mata, foram utilizados ganchos e pinças grandes para revirar o folhiço.
Confeccionou-se “sugadores” para extração de água de bromélias de grande porte, de suas axilas e tubo central. Toda a água retirada foi peneirada e devolvida para as mesmas plantas. Para bromélias muito pequenas utilizou-se pinça grande de 30cm; os detritos foram retirados, e as folhas afastadas para observação do seu interior. Algumas bromélias pequenas foram sacudidas em um tecido grande de cor clara, que evidencia melhor os organismos, e logo após eram devolvidas ao mesmo local.
Figura 63 - Localização dos pontos de coleta - herpetofauna (Legenda: 1 - Tarituba, riacho na mata de encosta; 2 - Tarituba, brejo na borda da mata; 3 - Mata da Usina; 4 - Mata da Marina; 5 - Bracuí, brejo; 6 - Piraquara de Fora; alagado na praia; 7 - Ilha de Pingo-d'Água; 8 - Mata da Usina; trilha do paiol; 9 - Rio Bracuí, mata às margens do rio).
Para a coleta dos girinos, utilizou-se puçá de três tamanhos básicos, pequeno, médio e grande com malhas mais finas e mais grossas.
Os adultos foram coletados manualmente através do método de observação direta e/ou através de sua vocalização. Ambos (girinos e adultos) foram transportados em sacos plásticos e acomodados em caixas de isopor, a fim de ficarem isolados da temperatura ambiente. O canto de algumas espécies foi registrado através de microfone direcional Sennheiser K 3U e gravador Sonny WM-D6C. A temperatura do ar e a umidade relativa foram registradas com um termômetro digital e um higrômetro, em todas as gravações de canto dos anfíbios. A temperatura e o pH da água foram medidos em alguns pontos de coleta dos girinos.
A busca do levantamento bibliográfico foi feita de três formas:
- no Zoological Records, em bibliotecas especializadas como a do Museu Nacional do Rio de Janeiro e Fundação Oswaldo Cruz (FIOCRUZ).
- trabalhos científicos feitos naquela região, onde foi utilizad o o programa Papyrus versão 7.0;
- trabalhos científicos envolvendo espécies possíveis de ocorrerem naquela região, onde se procuravam informações na lista do material, examinado pelos autores, para as localidades em questão.
O levantamento nas coleções herpetológicas foi feito através de busca para as localidades dos municípios de Parati, Angra dos Reis, Ilha Grande e Mangaratiba no estado do Rio de Janeiro.
No laboratório, o material de anfíbios recebido da coleta de campo foi anestesiado com cloretona 10% (adultos) e 5% (girinos), fixados em formol 10% (adultos) e 5% (girinos) e os adultos mantidos em álcool 70%. Posteriormente, foram triados e identificados com auxílio de microscópio estereoscópico Leica MZ6. O material de répteis foi anestesiado com éter sulfúrico, fixado em formol 10% e mantido em álcool 70%. Posteriormente para a identificação específica, recorreu-se a bibliografia especializada (Hoge & Romano-Hoge, 1978/79, Peters & Danoso-Barros, 1986). As vozes gravadas foram digitalizadas no programa Cool Edit Pro, para confirmação das espécies.
O trabalho de campo realizado resultou em dezoito espécies de anfíbios distribuídos em quatro famílias: Bufonidae (1), Centrolenidae (1), Hylidae (10) e Leptodactylidae (6) e três espécies de repteis. Os resultados foram sumariados, contendo as espécies encontradas, distribuídas pelos nove pontos de coleta, contendo ainda as informações quanto ao modo de coleta: Adulto (A), Filhote (F), Girino (G), Vocalização (V) e Observado (O). A seguir descrevemos os nove pontos de coleta para uma localização mais precisa, e apresentamos na Tabela 55 a distribuição das espécies encontradas.
- Ponto 1. Tarituba (incluindo o Morro de São Gonçalo), riacho dentro da mata de encosta.Ponto 2. Tarituba, brejo na borda da mata, na parte baixa.Ponto 3. Mata da Usina, riacho acima de Piraquara de Fora.Ponto 4. Mata da Marina, riacho ao nível do mar, próximo a Ponta da Pitanga.Ponto 5. Bracuí, brejo em mata de restinga, próximo as ruínas.Ponto 6. Lago em Piraquara de Fora, formado pelo riacho, com vegetação rasteira a arbustiva.Ponto 7. Ilha de Pingo d’Água, pequena ilha desabitada com vegetação arbustiva, bromélias, paredões rochosos e camada densa de folhiço.Ponto 8. Mata da Usina, trilha do paiol, vegetação bem preservada incluindo os estratos herbáceo, arbustivo e arbóreo.
- Ponto 9. Mata do rio Bracuí, parte alta (acima da BR-110), área com muitos sítios e algumas residências.
Tabela 55 - Espécies de anfíbios e répteis coletadas na excursão de maio de 2002, distribuídos através de seus respectivos pontos (P1 a P9).
| Espécie |
P1 |
P2 |
P3 |
P4 |
P5 |
P6 |
P7 |
P8 |
P9 |
| ANFÍBIOS |
|
|
|
|
|
|
|
|
|
| Adenomera marmorata |
|
|
A |
|
|
|
|
|
|
| Bufo crucifer |
|
|
|
G |
A |
|
|
|
A |
| Cycloramphus boraceiensis |
V |
|
|
|
|
|
|
|
|
| Eleutherodactylus binotattus |
A |
|
|
|
A |
|
|
A |
|
| Eleutherodactylus bolbodactylus bolbodactylus bolbodactylus |
|
|
|
|
|
|
|
V |
|
| Hyla albomarginata |
|
V |
|
|
|
|
|
|
|
| Hyla circumdata |
G |
|
|
|
|
|
|
|
|
| Hyla giesleri |
|
AV-V |
|
|
|
|
|
|
|
| Hyla minuta |
|
V |
|
|
|
|
|
|
|
| Hyla sp.aff albofrenata |
|
|
AGV |
|
|
|
|
|
|
| Hyalinobatrachium uranoscopum |
V |
|
|
|
|
|
|
|
|
| Hylodes phyllodes |
AG |
|
|
|
|
|
|
|
|
| Osteocephalus langsdorfii |
|
|
|
|
O |
|
|
|
|
| Scinax angrensis |
|
|
|
AV |
|
|
|
|
|
| Scinax humilis |
|
AV |
|
|
|
|
|
|
|
| Scinax hayii |
|
AV |
|
A |
|
V |
|
|
|
| Scinax perpusillus |
A |
|
V |
|
|
|
|
|
|
| Thoropa miliaris |
A |
|
AGF |
|
|
|
|
A |
|
| RÉPTEIS |
|
|
|
|
|
|
|
|
|
| Bothrops jararacussu |
F |
|
|
|
|
|
|
|
|
| Hemidactylus mabouia |
|
|
|
|
|
|
O |
|
|
| Tropidurus cf. torquatus |
|
|
|
|
|
|
O |
|
|
A= Adulto, G= Girino, V= Vocalização, O= Observado; F - Filhote
A mata na vizinhança imediata à usina Angra 1 é primária, com vegetação de Mata Atlântica (Veloso et al., 1991), bem preservada. Contudo nenhum espécime de réptil foi aí coletado. O relato de um dos guardas da Usina confirma a presença de lagartos (a descrição incerta não permite um parecer sobre a(s) espécies(s), de “cobra-coral” (família Elapidae ou Colubridae) e de “cobras verdes arborícolas” (provavelmente Chironius sp ou Philodryas olfersii, Colubridae). Nesta mata há uma trilha, já algo fechada, que se inicia próximo à casa da guarda e leva provavelmente até os fundos da Usina (não foi percorrida até o final). O caminho apresenta aspecto bem abandonado e com certeza é rara a circulação de pessoas por ali.
A vegetação próxima às praias e nas trilhas próximas à Usina Angra 1, em direção a Angra dos Reis, se apresentam bastante conservadas, apesar da proximidade da estrada e suas descidas em direção à praia e à Usina, em particular a que leva a um píer, regularmente freqüentada. Numa delas, foi encontrada uma casa abandonada e em ruínas. Estas regiões se apresentam como extensões da mata primária da Usina, e nelas foram coletados diversos anfíbios anuros.
Um pouco mais afastado da Usina Angra 1, do outro lado da estrada, fica o Morro de São Gonçalo, cujo sopé, pelo menos na vertente litorânea, apresenta vegetação já modificada pela presença de habitações e circulação humana. Na faixa dos 100 m. de altitude foi capturado um juvenil de Bothrops jararacussu (Viperidae).
A Ilha que melhor pôde ser observada foi a do Pingo d’Água, por sua pequena dimensão e proximidade do litoral. Ali foram observados um Gekkonidae (“lagartixa”; provavelmente Hemidactylus mabouia) e um Tropiduridae (“calango”, Tropidurus cf. torquatus). Na ilha ocorrem pelo menos quatro micro-habitais para répteis:
- paredões rochosos com vegetação esparsa;
- camada densa de folhiço;
- bromélias (em grande quantidade);
- áreas de vegetação arbustiva.
Há também pontos com cobertura arborícola pouco densa. Não há ocupação humana nesta ilha, nem, aparentemente, cursos d’água permanentes.
O número de espécies de anfíbios para a região totalizou em setenta e duas, de uma única ordem, Anura, distribuídas em seis famílias: Brachycephalidae (4), Bufonidae (3), Centrolenidae(2), Hylidae (35), Leptodactylidae (27) e Microhylidae (1). A Figura 64 ilustra estes resultados.
Figura 64 - Distribuição das Famílias de Anuros
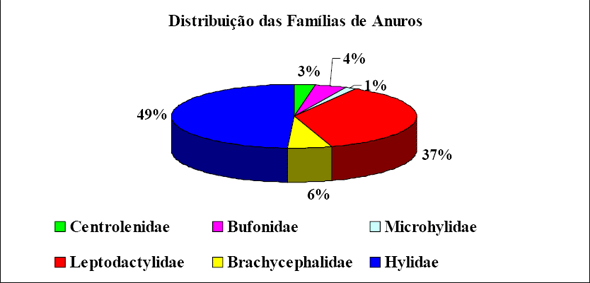
Tabela 56 - Espécies de anfíbios registradas em coleções herpetológicas e ou literatura para a região de influência da Usina de Angra dos Reis.
Clique aqui para visualizar a tabela 56
O número de espécies de répteis para a região totalizou 37, sendo uma da ordem Quelonia, subordem cryptodira e todas as outras da ordem squamata, das subordens: amphisbaenia (1), lacertilia (9) e serpentes (26). A Figura 65 ilustra estes resultados.
Figura 65 - Distribuição dos Grupos de Répteis
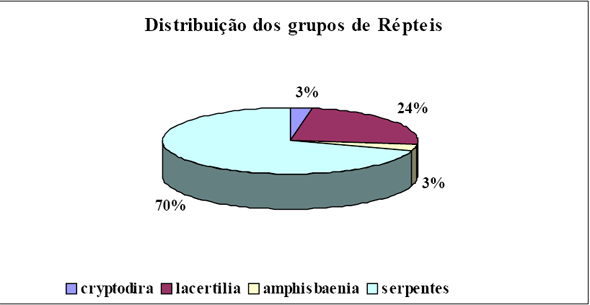
Tabela 57 - Espécies de répteis registradas em coleções herpetológicas e ou literatura para a região de influência da Usina de Angra dos Reis.
| Espécie |
Parati |
Angra |
Mangaratiba |
Ilha Grande |
Tarituba |
| Ameiva ameiva |
|
|
|
L |
|
| Boa constrictor |
|
|
|
C-L |
|
| Bothrops jararaca |
|
C |
L |
L |
|
| Bothrops jararacussu |
C |
|
|
L |
C |
| Bothrops sp |
|
|
|
L |
|
| Chironius bicarinatus |
|
|
L |
L |
|
| Chironius fuscus |
|
|
C-L |
|
|
| Corallus hortulanus |
|
|
|
C-L |
|
| Dipsas albifrons |
|
|
|
L |
|
| Ecpleopus gaudichaudii |
|
|
L |
|
|
| Enyalius brasiliensis |
|
|
L |
|
|
| Epicrates cenchria |
|
|
|
L |
|
| Erythrolamprus aesculapii |
|
|
L |
|
|
| Geochelone carbonaria |
|
|
|
L |
|
| Gymnodactylus geckoides |
|
|
L |
|
|
| Hemidactylus mabouia |
|
L |
|
|
|
| Leposternon microcephalum |
|
|
L |
|
|
| Liophis miliaris |
|
|
|
L |
|
| Liophis ["Leimadophis"] poecilogyrus |
|
|
L |
L |
|
| Mastigodryas bifossatus |
|
|
|
L |
|
| Micrurus corallinus |
|
|
L |
C-L |
|
| Oxyrhopus clathratus |
|
|
L |
|
|
| Philodryas olfersii |
|
|
L |
L |
|
| Philodryas serra |
|
|
L |
|
|
| Pseudoboa cloelia |
|
|
|
L |
|
| Sibynomorphus newviedi |
|
|
L |
C |
|
| Sibynomorphus turgidus |
|
|
|
L |
|
| Siphlophis pulcher |
|
|
L |
|
|
| Spilotes pullatus |
|
|
L |
L, C |
|
| Thamnodynastes pallidus |
|
|
|
L |
|
| Thamnodynastes strigilis |
|
|
L |
|
|
| Tropidurus torquatus |
|
|
|
L |
|
| Tupinambis merianae |
|
|
|
L |
|
| Tupinambis teguixin |
|
|
|
L |
|
| Xenodon neuwiedii |
|
|
L |
|
|
OBS.: L -Literatura; C - Coleção.
A Mata Atlântica é considerada um dos maiores centros de biodiversidade do planeta, com alto grau de endemismo. Segundo Lynch (1979) 92% das 183 espécies de anfíbios que ocorrem na Mata Atlântica são endêmicas, tendo inclusive uma família (Brachycephalidae) e uma sub-familia (Elosiinae) da família Leptodactylidae, endêmicas. Na região da CNAAA a família Brachycephalidae está representada por quatro espécies dos dois únicos gêneros, sendo duas destas restritas para esta região: Brachycephalus vertebralis e Psyllophryne hermogenesi. A sub-familia Elosiinae, na região em estudo está representado por 4 espécies de dois gêneros: Megaelosia goeldii, Megaelosia bocainesis, Hylodes phyllodes e Hylodes asper Atualmente, segundo Frost (2002), para a Mata Atlântica o número total de espécies de anfíbios é de 495, registrando um aumento de 270% de espécies em 23 anos. Na região de estudo, por exemplo, encontrou-se uma espécie nova de cor verde do gênero Hyla do grupo Albofrenata pertencente a família Hylidae, que está sendo descrita. Assim como esta, outras espécies inéditas devem estar presentes na região, que ainda não foi suficientemente estudada.
As listas oficiais da fauna ameaçada de extinção dos estados do Rio de Janeiro e de São Paulo utilizaram para classificar os status de conservação das espécies, as normas da IUCN, como por exemplo: (EP) em perigo e (PA) presumivelmente ameaçadas. Pelo menos seis espécies com distribuição para a região constam destas listas: Paratelmatobius gaigeai (EP), Flectonotus ohousi (PA) Maegaelosia bocainensis (PA), Cyclorhamphus eleutherodactylus (PA), Hyalinobatrachium uranoscopum (PA) e Hyla clepsydra (PA).
Hyla clepsydra é uma espécie rara de distribuição restrita, provavelmente com população pouco abundante e está registrada, na região, em coleções científicas, presentemente consultadas.
Hyalinobatrachium uranoscopum é uma espécie sensível, dependente de matas protegidas, que se reproduz nas folhas de arbustos acima do nível dos riachos, e que foi localizada no ponto dois de coleta (P2).
Flectonotus ohousi é uma espécie com hábitos muito peculiares de reprodução, seus girinos são criados em água acumulada nos orifícios do bambú.Está assinalada para as serras de São Paulo e Rio de Janeiro, tendo sido encontrada em Mangaratiba, RJ, devendo portanto, ocorrer na área estudada.
O ciclo vital da maioria dos anfíbios envolve uma larva aquática e um adulto terrestre. Esta particularidade faz com que os impactos ambientais que afetam os diversos tipos de ambientes aquáticos, nos quais os girinos se desenvolvem, ou aqueles que atinjam ambientes terrestres, sejam igualmente refletidos nas densidades populacionais deste animais. A permeabilidade do tegumento, por outro lado, pode tornar os anuros particularmente susceptíveis às alterações em parâmetros físico-químicos do ambiente(Caramaschi et al. 2000). Esta sensibilidade aliada ao ciclo de vida e as especializações reprodutivas conferem aos anfíbios, a capacidade de serem excelentes indicadores de qualidade ambiental (bioindicadores).
A região onde está localizada a CNAAA, representa uma área de concentração e reprodução de muitas espécies de anfíbios e répteis. Constatou-se a presença de uma mata recuperada em seu entorno, com boa vegetação e diversos micro-ambientes propícios para a herpetofauna. O número de espécies encontradas para os anfíbios é muito superior ao registrado no Plano de Manejo da Estação Ecológica de Tamoios (Fase 1), três gêneros sem identificação. Este fato se deve porque muitos dados importantes sobre a composição de fauna brasileira não têm sido publicados, principalmente por falta de incentivo, estando as informações limitadas a monografias, dissertações, teses e relatórios, que muitas vezes são de difícil acesso. Corroborando com Haddad, 1998, estudos de composição faunística são fundamentais para a compreensão da biodiversidade e consequentemente para o planejamento de tomada de decisões sobre estratégias de conservação.
Do total de anfíbios obtidos na área de estudo, 72 espécies acredita-se que somente cerca de 16%, restritas das partes altas da Serra da Bocaina, não devam sofrer influência da Usina.
Das três ilhas visitadas a Pingo d’Água foi que apresentou ambientes mais propícios a existência de anfíbios e repteis.
O período reprodutivo dos anfíbios é a época em que ficam mais expostos, principalmente pela emissão de voz, o que permite uma avaliação mais acertada da população. Para a maioria das espécies, o período reprodutivo é de setembro a início de novembro.
Mesmo com um número reduzido de espécies obtidas no período da coleta, pôde ser feita uma avaliação da herpetofauna da região de Angra dos Reis e proximidades, pois desde há algum tempo, tem sido feito pelos presentes autores um trabalho de pesquisa nas localidades de Parati, Angra dos Reis, Ariró, Iha Grande, Mangaratiba e Tarituba, no Estado do Rio de Janeiro, e Serra da Bocaina, no Estado de São Paulo. Os dados de coleta destes trabalhos estão tombados principalmente nas coleções herpetológicas da UNIRIO e UFRJ.
As observações de campo evidenciaram variações de ambiente nos locais visitados, e, como foi citado, por exemplo, para a Ilha do Pingo d’Água e para a mata vizinha à Usina Angra 1, há diferentes micro-habitats favoráveis para a ocorrência de espécies de répteis.
A coleta de diferentes espécies de anfíbios anuros também aponta para uma provável presença de variadas espécies de répteis, uma vez que são presas potenciais (e.g. para Tupinambis spp, Chironius bicarinatus, C. fuscus, Liophis miliaris, L. poecilogyrus, Philodryas olfersii, Thamnodynastes spp, Xenodon neuwiedii, Bothrops jararaca juvenis, B. jararacussu - todas espécies citadas para a área em relatório anterior). Desta forma, pode-se dizer que a região visitada, como um todo, comporta herpetofauna bem representativa. Averiguações são necessárias sob mais de um aspecto:
-
em relação à ocorrência mesma de várias espécies, citadas em bibliografia (cf. relatório anterior) mas não capturadas ou observadas nesta campanha. Assim, Sibynomorphus turgidus, Thamnodynastes pallidus nattereri, T. strigilis e a “urutu” (Botrhops sp) necessitam de verificação de ocorrência e identificação. Além disso, muitas referências são de áreas próximas, mas não diretamente afetadas pelo projeto;
-
a veracidade dos relatos de alguns moradores, não em relação à ocorrência, mas sim à identidade específica dos animais citados, como é o caso das “caninanas” e “cobras-corais” referenciadas para as ilhas;
-
ampliação dos registros já obtidos, pois as coletas da presente campanha não acrescentaram novidades à listagem anterior de espécies
Apesar destas dificuldades, o prognóstico é muito favorável em relação à riqueza da herpetofauna, e assim, por exemplo, de acordo com os relatos obtidos, na área próxima à Usina Angra 1 devem ocorrer Chironius spp e Philodryas olfersii, bem como a coral-verdadeira Micrurus corallinus) e falsa (provavelmente Erytrolamprus aesculapii e Oxyrhopus spp). Nas áreas de folhiço, há boa probabilidade do encontro de lagartos das famílias Scincidae (Mabuya spp) e Gymnophtalmidae (“microteídeos”), que eventualmente podem estar representadas por espécies raras. Isto significa, por um lado, que a presença de espécies mais comuns não está afetada, e por outro que existe ainda a possibilidade do encontro de espécies pouco observadas ou coletadas.
Durante o período de navegação, foi feita a tentativa de localização de quelônios marinhos. Apesar de não terem sido observadas diretamente, sabe-se que a região é freqüentada por tartarugas (Chelonia mydas, Caretta caretta, Lepidochelys olivacea, Eretmochelys imbricata, Dermochelys coriacea; Márquez, 1977, Friberg, 1981).
A presença de serpentes peçonhentas (Bothrops, Micrurus, Philodryas olfersii) levanta sempre a questão de riscos com acidentes ofídicos. Porém, nada foi apurado em relação a algum caso de envenenamento, supostamente não usual para a região, pois as espécies em questão são encontradas apenas dentro da mata, não chegando habitualmente a áreas de moradia humana. Constatou-se que a área do entorno imediato à CNAAA está bem preservada, enquanto que a região de Angra dos Reis sofre um acelerado processo de degradação ambiental por desmatamento. Esta situação é devida, certamente, ao respeito da população com relação à área da Usina, e ao fato de que sobre esta se mantém uma vigilância.
Espécies de anfíbios observadas na área de influência da CNAAA (maio de 2002)
Adenomera marmorata Steindachner, 1867. - Pequena rã que vive no chão da mata e capinzais. Para se reproduzir, escavam pequeno túnel que termina em uma câmara subterrânea, onde o casal faz um ninho de espuma onde deposita seus ovos. A espécie é assinalada desde o Estado do Rio de Janeiro até o Estado de Santa Catarina.
Bufo crucifer Wied-Neuwied, 1821.- Espécie que habita áreas abertas apresentando glândulas paratóides. Desovam preferencialmente em poças de água parada. São encontrados em grande parte do território brasileiro (Izecksohn & Carvalho e Silva, 2001). Suas larvas de coloração negra aglomeram-se em grandes quantidades nas margens de rios e nos lagos onde os adultos reproduzem. Os girinos foram encontrados em rio próximo a marina, próximo a Ponta da Pitanga (Ponto 4).
Cycloramphus boraceiensis Heyer, 1983. - rã encontrada na Serra do Mar, nos Estados do Rio de Janeiro e São Paulo, na Ilha Grande e na Ilha de São Sebastião (Frost, 2002).
Eleutherodactylus binotatus (Spix, 1824).- Rã de porte médio que apresenta desenvolvimento direto,. É uma espécie estritamente florestal que vive no chão da mata e está distribuída pelo Sudeste brasileiro (Izecksohn & Carvalho e Silva, 2001). Registrada nas coleções ZUFRJ e UNIRIO para os municípios de Mangaratiba e Angra dos Reis, RJ. Encontrada no folhiço nos pontos 1, Tarituba, RJ, ponto 5, Angra dos Reis e ponto 8, Bracuí, RJ.
Eleutherodactylus bolbodactylus (Lutz, 1925). -Rã que vive em folhiço, descrita do município de Angra dos Reis, sendo que recentemente foi redescrita da mesma localidade (Pombal & Cruz, 1999).Sua voz foi registrada no ponto 8, próximo a Usina, Angra dos Reis, RJ.
Hyla albomarginata Spix, 1824.- Perereca verde, de porte médio, que habita áreas abertas ou bordas de mata e que vocaliza na margem dos brejos e lagos. Trata-se de uma espécie de distribuição ampla, incluindo países vizinhos (Izecksohn & Carvalho e Silva, 2001). Registrada nas coleções ZUFRJ e UNIRIO para os municípios de Mangaratiba e Angra dos Reis, RJ.
Hyla aff. albofrenata - Espécie nova de perereca verde, encontrada na região, pertencente ao grupo “albofrenata”, que está sendo descrita. Registrada nas coleções ZUFRJ e UNIRIO para os municípios de Mangaratiba e Angra dos Reis, RJ. Foram encontrados além de girinos, machos vocalizando e fêmea a poucos centímetros da água no ponto 3.
Hyla circumdata (Cope, 1871).- Perereca de grande porte, de cor marrom, que habita regiões alagadas no interior da mata. Registrada nas coleções ZUFRJ e UNIRIO para os municípios de Mangaratiba e Angra dos Reis, RJ. Foram encontrados girinos desta espécie em Tarituba, no ponto 1.
Hyla giesleri Mertens, 1950.- Perereca encontrada na região litorânea de Mata Atlântica, entre os Estados do Espírito Santo e Rio de Janeiro (FROST, 2002). Habita áreas alagadas e lagos em áreas abertas. Registrada nas coleções ZUFRJ e UNIRIO para os municípios de Mangaratiba e Angra dos Reis, RJ. Encontrada vocalizando em poça no ponto 2.
Hyla minuta Peters, 1872.- Perereca que canta na vegetação baixa de brejos e poças em áreas abertas. Os ovos são postos na superfície da água (Izecksohn & Carvalho E Silva, 2001). Registrada nas coleções ZUFRJ e UNIRIO para os municípios de Mangaratiba e Angra dos Reis, RJ. Encontrada vocalizando em poça no ponto 2.
Hyalinobatrachium uranoscopum (Müller, 1924). Pequena perereca com colorido verde e ventre transparente, com focinho espatulado e voz semelhante a insetos noturnos. Habita arbustos, onde deposita seus ovos, nas margens dos córregos e ribeirões das matas de encosta (Izecksohn & Carvalho e Silva, 2001). Seu canto agudo foi identificado por nós no início da trilha no ponto 1, em Tarituba, RJ, nas margens do riacho, em noite chuvosa.
Hylodes phyllodes Heyer and Cocroft, 1986. - Espécie diurna que vive e cria suas larvas em riachos encachoeirados no interior da Mata . Registrada para os municípios de Angra dos Reis na Ilha Grande, (ROCHA et al, 1997) e no município de Mangaratiba, RJ nas coleções de anfíbios ZUFRJ e UNIRIO. Sua voz característica lembra o canto de aves. Foi encontrada vocalizando no riacho em Tarituba, RJ, durante o dia, no ponto 1.
Osteocephalus langsdorffii (Duméril and Bibron, 1841) - Perereca de grande porte, especialmente a fêmea, que pode ultrapassar 10 cm de comprimento, do focinho ao ânus. Os machos vocalizam dilatando um duplo saco vocal. A espécie tem distribuição ampla podendo se afastar muito do litoral (Izecksohn & Carvalho e Silva, 2001). Registrada na coleção UNIRIO para o município de Mangaratiba, RJ. Encontrada por nós em brejo de restinga em Bracuí, RJ, no ponto 5.
Scinax angrensis (Lutz, 1973)- Espécie que desova e cria suas larvas em remansos de riacho em matas de encosta (Carvalho e Silva et al, 1995). Estes autores examinaram material proveniente de Mangaratiba e Parati, estando presente nas coleções ZUFRJ e UNIRIO nos dois municípios. Foi encontrado o macho vocalizando em riacho, nas imediações da Usina, no ponto 4.
Scinax hayii (Barbour, 1909).- Espécie de ampla distribuição pelas serras do Sudeste brasileiro. Procria em lagos e alagados, e também em reservatórios artificiais, desde que com alguma vegetação (Bokermann, 1967). Registrada nas coleções ZUFRJ e UNIRIO para os municípios de Mangaratiba e Angra dos Reis, RJ. Encontrada vocalizando nos pontos 2, 4 e 6, nas imediações da Usina, Angra dos Reis, RJ.
Scinax humilis (B. Lutz, 1954) - Espécie de mata de baixada, em áreas onde ocorre a formação de poças no chão da mata (Carvalho e Silva & Carvalho e Silva, 1998). Tendo sido registrada na coleção herpetológica ZUFRJ para Parati, RJ. Foi encontrada vocalizando em brejo, no ponto 2.
Scinax perpusillus (A. Lutz and B. Lutz, 1939) - Espécie que ocorre em formações de restinga, em formações de broméliaceas instaladas sobre paredões rochosos litorâneos, e em áreas de mata de encosta ou baixadas (Peixoto, 1998). O autor examinou exemplares provenientes da Ilha Grande, RJ. Registrada nas coleções ZUFRJ e UNIRIO para os municípios de Mangaratiba e Angra dos Reis, RJ. Encontrada vocalizando em bromélias no ponto 1, em Tarituba, RJ e no ponto 3, nas imediações da Usina, Angra dos Reis, RJ.
Thoropa miliaris (Spix, 1824) - Rã de porte médio, adaptada para viver em paredões rochosos onde correm filetes de água. Sua biologia reprodutiva foi estudada por Oliveira (1999), na região da Urca (Rio de Janeiro, RJ) e de Mangaratiba, RJ. Registrada nas coleções ZUFRJ e UNIRIO para os municípios de Mangaratiba e Angra dos Reis, RJ. Encontrados adultos, filhotes e girinos e no ponto 1 e em Angra, próximo ao paredão de saída de água quente da Usina e adultos no ponto 3 em Tarituba, RJ.
Espécies de anfíbios na área de influência da usina de Angra
Brachycephalus ephippium (Spix, 1824)- Pequeno sapo de cor amarela endêmico da Mata Atlântica, assinalado para a região de Angra dos Reis e Mangaratiba, RJ.
Brachycephalus vertebralis (Pombal, 2001) - Espécie descrita recentemente para o município de Parati, RJ
Bufo ictericus Spix, 1824- Espécie de grande porte que habita áreas abertas, apresentando glândulas paratóides. Desovam preferencialmente em poças de água parada.. São encontrados em grande parte do território brasileiro (Izecksohn & Carvalho e Silva, 2001).
Crossodactylus gaudichaudii Duméril and Bibron, 1841 - Rã de tamanho médio e hábitos diurnos, que vive sobre as pedras dos córregos. Desova sob as pedras nos córregos, e girinos se desenvolvem nesse ambiente. Ocorre no Sudeste brasileiro (Izecksohn & Carvalho e Silva, 2001).
Cycloramphus eleutherodactylus (Miranda-Ribeiro, 1920) -Rã de tamanho médio, com dedos e artelhos sem membranas, focinho arredondado e estritamente florestal (Izecksohn & Carvalho e Silva, 2001).
Cycloramphus fulginosus Tschudi, 1838 - Rã de tamanho grande e corpo deprimido, com os pés palmados e colorido com aspecto de fuligem. Vive entre ou sobre pedras nos córregos das encostas das florestas (Izecksohn & Carvalho e Silva, 2001). Registrada para Ilha Grande, na coleção UNIRIO.
Cycloramphus granulosus Lutz, 1929 - Rã de tamanho médio e colorido marrom escuro, que possui os pés posteriores não palmados e dorso densamente coberto por grânulos. Ocorre em Bonito, na Serra da Bocaina (Lutz, 1929).
Dendrophryniscus brevipollicatus Jiménez de la Espada, 1871 - Pequeno bufonídeo desprovido de glândulas paratóides, que cria suas larvas em bromélias (Izecksohn, 1993). Este autor examinou material proveniente de Angra dos Reis, Mangaratiba e Parati, RJ e Serra da Bocaina, SP. Registrada nas coleções ZUFRJ e UNIRIO para os municípios de Mangaratiba e Ubatuba, SP.
Eleutherodactylus guentheri (Steindachner, 1864) - Rã de porte médio, com o primeiro e segundo dedos de igual tamanho, sem o par de pintas dorsais e com uma estria enegrecida ao longo da face externa das tíbias. Vive no chão da mata onde depositam seus ovos. O desenvolvimento é direto, nascendo rãzinhas já metamorfoseadas (Izecksohn & Carvalho e Silva, 2001).
Eleutherodactylus nasutus (A. Lutz, 1925) - Espécie de montanha, que também possui desenvolvimento direto, sem estágio larval. Possui um focinho longo e habita áreas abertas, sob a serapilheira (Lynn & Lutz, 1946).
Eleutherodactylus parvus (Girard, 1853) - Pequena rã diurna que vive em folhiço de mata.
Eleutherodactylus pusillus Bokermann, 1967 - Pequena rã que vive no folhiço da mata, sendo descrita de regiões da Serra da Bocaina a partir de 1600 m.
Flectonotus goeldii (Boulenger, 1895)- Perereca bromelígena que transporta seus ovos no dorso, de onde nascem girinos que completam seu desenvolvimento na água do interior das bromélias. (Izecksohn & Carvalho e Silva, 2001).
Flectonotus ohausi (Wandolleck, 1907)- Perereca que habita bambuzais, transportando seus ovos no dorso, de onde nascem girinos que completam seu desenvolvimento na água acumulada no interior dos bambus.
Gastrotheca albolineata (A. Lutz and B. Lutz, 1939)- Perereca verde que habita bromélias epífitas. Apresenta um “marsúpio dorsal” onde carrega seus ovos até a eclosão, onde dá origem a pequenos sapos.
Hyalinobatrachium eurygnathum (A. Lutz, 1925) - Pequena perereca com colorido verde e ventre transparente, com focinho curto e voz semelhante a insetos noturnos. Habita arbustos, onde deposita seus ovos, nas margens dos córregos e ribeirões das matas de encosta (Izecksohn & Carvalho e Silva, 2001).
Hyla anceps A. Lutz, 1929 - Perereca de porte médio, com colorido do dorso marrom, ventre vermelho e anéis negros nas coxas. Reproduz-se em poças, de preferência com alguma profundidade, onde se desenvolvem suas larvas. Distribui-se pelos Estados do Rio de Janeiro, Minas Gerais, Espírito Santo e sul da Bahia (Izecksohn & Carvalho e Silva, 2001).
Hyla arildae Cruz and Peixoto, 1985 - Espécie de perereca de cor verde, que cria suas larvas em riachos de Mata Atlântica. Registrada nas coleções ZUFRJ e UNIRIO para a Serra da Bocaina, SP.
Hyla bipunctata Spix, 1824 - Perereca que habita e reproduz em poças de áreas abertas em bordas ou não de matas, onde deposita suas desovas diretamente na água. Apresenta um canto característico que lhe dá o nome vulgar de “sapo-telégrafo”. Registrada nas coleções ZUFRJ e UNIRIO para Mangaratiba e Tarituba, RJ.
Hyla callipygia (Cruz & Peixoto, 1984) - Perereca de cor verde, descrita da Serra da Bocaina, SP.
Hyla clepsydra Lutz, 1925 - Perereca verde descrita da Serra da Bocaina e citada por Bokermann (1972) para o Parque Nacional da Serra da Bocaina. Há exemplares tombados na coleção herpetológica EI da Estrada Parati-Cunha.
Hyla decipiens A. Lutz, 1925 - Perereca que habita e reproduz em poças de áreas abertas em bordas ou não de matas, onde deposita suas desovas na vegetação acima d’água. Apresenta vistosa moldura clara em torno do corpo. Registrada na coleção UNIRIO para o município de Mangaratiba, RJ.
Hyla elegans Wied-Neuwied, 1824 - Perereca que habita e reproduz em poças de áreas abertas em bordas ou não de matas, onde deposita suas desovas diretamente na água. Apresenta vistosa moldura clara em torno do corpo e membros. Registrada nas coleções ZUFRJ e UNIRIO para os municípios de Mangaratiba e Angra dos Reis, RJ.
Hyla faber Wied-Neuwied, 1821 - Perereca de grande porte que habita e reproduz em poças de áreas abertas em bordas ou não de matas, onde deposita suas desovas em ninhos ou “panelas” nas bordas das poças. Apresenta um canto característico que lembra “o bater de um martelo em metal”, por isso é vulgarmente chamado de “sapo-ferreiro”, “sapo-martelo” ou “sapo-bigorna”. Registrada nas coleções ZUFRJ e UNIRIO para os municípios de Mangaratiba e Angra dos Reis, RJ.
Hyla senicula Cope, 1868 - perereca de porte médio, com padrão dorsal e colorido simulando casca de árvores. Utiliza poças nas clareiras ou bordas de mata para reproduzir e desovam, provavelmente, na superfície da água (Izecksohn & Carvalho e Silva, 2001).
Hyla semilineata Spix, 1824 - Perereca de cor marrom, que habita e reproduz em poças de áreas abertas, onde deposita sua desova na água. Os girinos de coloração negra formam cardumes que nadam na superfície d’água. Registrada nas coleções ZUFRJ e UNIRIO para os municípios de Mangaratiba e Angra dos Reis, RJ.
Hylodes asper. (Müller, 1924) - Espécie diurna que vive e cria suas larvas em riachos encachoeirados no interior da Mata. Tendo sido registrada nas coleções de anfíbios ZUFRJ e UNIRIO do município de Mangaratiba, RJ. Sua voz característica lembra o canto de aves.
Leptodactylus ocellatus (Linnaeus, 1758) - Rã de grande porte, especialmente o macho, que podem apresentar braços muito espessos. É uma espécie que resiste a alterações ambientais antrópicas e cujos girinos parecem suportar um grau de poluição da água não aceitável por outras espécies de anuros. Desovam na superfície da água onde fazem ninhos de espuma (Izecksohn & Carvalho e Silva, 2001).
Leptodactylus spixii Heyer, 1983 - Rã de tamanho médio, com cordões cutâneos longitudinais dorsais e estria enegrecida sobre lábio superior. Habita florestas e suas bordas. Escava câmaras subterrâneas, ao redor de brejos ou poças dentro das quais o macho vocaliza e onde realizam a desova, também em ninhos de espuma (Izecksohn & Carvalho e Silva, 2001). Registrada na coleção UNIRIO para o município de Mangaratiba, RJ.
Megaelosia goeldii (Baumann, 1912) - Rã diurna de grande porte, com girino de grandes dimensões, que vive em riachos de matas em regiões montanhosas da Serra dos Örgãos e Friburgo, RJ. Também registrada para a região de Parati, RJ na coleção ZUFRJ.
Megaelosia bocainensis (Giaretta et al., 1993) - Rã diurna de grande porte, que vive em riachos da Serra da Bocaina.
Myersiella microps Carvalho, 1954 - Rã de pequeno porte, com corpo globoso, cabeça muito pequena e focinho afilado. Vive no chão das florestas úmidas. Possui desenvolvimento direto, tendo, porém, embriões com forma de girino dentro do corpo. Habitam as florestas da Serra do Mar, nos Estados do Rio de Janeiro e de São Paulo (Izecksohn & Carvalho e Silva, 2001). Foi registrada na coleção UNIRIO na Ilha Grande, RJ.
Paratelmatobius gaigae (Cochran, 1938) - Pequeno sapo, habitante do chão de matas preservadas da Serra da Bocaina a partir de 1100 m de altitude.
Phasmahyla guttata (A. Lutz, 1925) - Perereca que habita e reproduz em riachos encachoeirados no interior de matas. Pertence a um grupo que apresenta a pupila ocular vertical. Sua devosa é posta na vegetação acima d’água, de onde originam girinos que nadam em cardumes na correnteza destes riachos. Registrada nas coleções ZUFRJ e UNIRIO para os municípios de Mangaratiba e Angra dos Reis, RJ.
Physalaemus barrioi (Bokermann, 1967a) - Pequena rã, habitante da Serra da Bocaina à aproximadamente 1600 m. reproduz em áreas abertas, sob a vegetação, principalmente após chuvas intensas.
Physalaemus moreirae (Miranda-Ribeiro, 1937) - Rã de porte pequeno e que ocorre Sudeste do Brasil (Caramaschi & Caramaschi, 1991).
Physalaemus signifer (Girard, 1853) - Rã de tamanho pequeno, com dorso escuro constituído por duas pontas de flecha. A região inguinal é, freqüentemente, de cor salmão e tem, de cada lado, uma glândula negra. Faz ninhos de espuma sob folhas mortas úmidas sobre a lama (Izecksohn & Carvalho e Silva, 2001). Registrada na coleção UNIRIO para Ilha Grande, RJ.
Phrynohyas mesophaea (Hensel, 1867) - Perereca grande, de cor bege, com dorso marrom-escuro e uma moldura branca ou bege-clara. Se oculta nas bromélias ou nas bananeiras, nas bordas ou na clareiras das florestas (Izecksohn & Carvalho e Silva, 2001).
Phrynomedusa marginata (Izecksohn and Cruz, 1976) - Espécie pequena, com o dorso e o ventre finamente granulado, sendo maiores sob a metade basal das coxas e na margem do ânus. Seu canto pode ser interpretado como um som débil, não muito grave, pulsado e de curta duração (Izecksohn & Cruz, 1976).
Proceratophrys appendiculata (Günther, 1873) - Ocorre nas vertentes voltadas para o litoral na Serra do Mar, nos Estados do Rio de Janeiro, São Paulo e Paraná (Izecksohn, 1998, Izecksohn et al, 1998), tendo sido por nós coletado nos Municípios de Parati e Mangaratiba, RJ.
Proceratophrys boiei (Wied-Neuwied, 1824). - Rã de porte grande, com aspecto de sapo, que se distingue pelos apêndices cutâneos nas pálpebras superiores, parecendo chifres. Reproduz-se em poças e remansos de riachos, com água renovável. Ampla distribuição pela Mata Atlântica (Izecksohn & Carvalho e Silva, 2001).
Psyllophryne didactyla Izecksohn (1971)- Menor vertebrado terrestre, vive em folhiço, que segundo Frost (1985), é endêmico da Mata Atlântica do Sudeste do Brasil. Tendo sido registrada na coleção UNIRIO para o município de Mangaratiba, RJ e citado município de Angra dos Reis em Ilha Grande.
Psillophryne hermogenesi (Giaretta E Sawaya, 1998) - Pequeno anuro encontrado em folhiço, endêmico da Mata Atlântica do Sudeste do Brasil. Sua distribuição inclui Ubatuba, SP, e adjacências de Parati, RJ.
Scinax alter (B. Lutz, 1973) - Perereca pequena, muito comum nos brejos da baixada aberta, onde desovam e criam suas larvas (Izecksohn & Carvalho e Silva, 2001).
Scinax ariadne (Bokermann, 1967) - Encontra-se na Serra da Bocaina, no Sudeste Brasileiro (Frost, 2002).
Scinax argyreornatus (Miranda-Ribeiro, 1926) - Os adultos dessa espécie são frequentemente encontrados próximos a poças dentro da mata, e durante o dia, podem ser encontrados em bromeliáceas e folhas enroladas, ou quando a mata está úmida, em topos de folhas de arbustos (Carvalho e Silva & Carvalho e Silva, 1998). Tendo sido registrada na coleção herpetológica ZUFRJ para Mangaratiba e Ariró.
Scinax atratus (Peixoto 1988) - Espécie que vive em bromélia, descrita de São José do Barreiro, Serra da Bocaina, SP.
Scinax cuspidatus (A. Lutz, 1925) - Perereca pequena cuja extremidade do focinho é acuminada. Espécie bromelícola obrigatória, mas reproduz-se em brejos, nas bordas das florestas ou próximos a estas. Os ovos são postos na superfície da água, onde as larvas se desenvolvem (Izecksohn & Carvalho e Silva, 2001).
Scinax eurydice (Bokermann, 1968) - Espécie do grupo “rubra”, e que distribui-se desde o Sul do Estado da Bahia até o Estado do Rio de Janeiro (Frost, 2002).
Scinax fuscovarius (A. Lutz, 1925) - Perereca pertencente ao grupo “rubra”, encontrada no Sudeste do Brasil, entre 150-1800m (Frost, 2002).
Scinax trapicheiroi (B. Lutz, 1954) - De acordo com Carvalho e Silva & Carvalho e Silva (1994), essa espécie vive em matas de encosta de serras litorâneas como Maciço da Tijuca, Pedra Branca e Ilha Grande, RJ. Registrada nas coleções ZUFRJ e UNIRIO para o município de Angra dos Reis, Ilha Grande, RJ.
Sphaenorhincus orophilus (A. Lutz and B. Lutz, 1938) - Pequena rã verde descrita de Petrópolis, RJ, tendo ainda, como distribuição, segundo Cruz & Peixoto (1979), outras localidades, como Bonito, na Serra da Bocaina, na divisa com o Estado de São Paulo.
Zachaenus parvulus (Girard, 1853) - Van Sluys et al. (2001) estudaram uma população desse pequeno sapo, que vive no folhiço, na Ilha Grande, RJ.
7.3.2. Avifauna - topo
Atualmente, a biodiversidade é um dos principais elementos utilizados para caracterizar a importância de uma área em termos de conservação. No Brasil, as aves têm sido empregadas como um dos principais veículos na tomada de decisões nesse sentido. Comparadas a outros grupos zoológicos, elas são de fácil amostragem, alta detectabilidade, além de serem o grupo de vertebrados mais conhecido na América do Sul. O conhecimento da avifauna local nos fornece ainda mecanismos para monitorar eventuais distúrbios sofridos pelo meio-ambiente. Assim, o desaparecimento de uma espécie, ou mesmo a colonização por uma espécie exótica, podem indicar alterações ambientais significativas.
Até meados da década passada, o conhecimento da avifauna do litoral sul do Estado do Rio de Janeiro restringia-se a informações provenientes de expedições de coleta empreendidas na primeira metade do século por Olivério Pinto, Herbert Berla e Lauro Travassos, além de material coletado por Helmut Sick na Ilha Grande. A maior parte desse material está depositada no Museu Nacional e no Museu de Zoologia da Universidade de São Paulo, mas não constava da literatura ornitológica até poucos anos atrás. Apenas recentemente, após o inventário dessas coleções ter sido feito em Pacheco et al. (1997), parte das informações foi apresentada ao meio científico. Esses autores apresentam ainda dados sobre aves coletadas na região e depositadas na Universidade Federal Rural do Rio de Janeiro. Todo esse material serve como registro histórico da avifauna local, apresentando inclusive informações sobre espécies atualmente muito raras ou consideradas extintas em todo o litoral sul do Estado do Rio de Janeiro. Pacheco et al. (1997) incluem ainda uma vasta lista de aves observadas na região pelos autores no período de 1983 a 1997, mas que ainda não haviam sido citadas na literatura, acrescentando 169 registros inéditos.
Outra fonte recente de informações de grande relevância é o inventário da avifauna relativo ao Programa de Gestão da Baía da Ilha Grande realizado por Dante Buzzetti em 1996 e 1997. Além desses dados, o autor apresentou uma lista de 333 espécies com as respectivas localidades e altitudes de registro em um artigo sobre a distribuição altitudinal das aves nos municípios de Angra dos Reis e Parati (Buzzetti, 1998).
Igualmente importantes são os registros de quase 300 espécies de aves nas baixadas da Costa Verde feitos por Elmiro Mendonça no período de 1997 a 2000 durante o seu trabalho com o papa-formigas-de-cabeça-negra Formicivora erythronotos, pássaro ameaçado de extinção endêmico da região. Esses dados tornam-se particularmente relevantes por terem sido obtidos em sua grande maioria no vale do Rio Mambucaba, região próxima às futuras instalações de Angra 3. Deve-se, ainda, considerar que a continuidade dos trabalhos realizados por E. Mendonça permitiu a obtenção de interessantes dados sobre migrações altitudinais e latitudinais da avifauna local.
7.3.2.1. Caracterização da Avifauna Terrestre - topo
Estudos com aves insulares também contribuem para o conhecimento da avifauna terrestre da região, com destaque para os trabalhos realizados na Ilha Grande. Além dos referidos trabalhos de Pacheco et al. (1997) e Buzzetti (1998), vários outros pesquisadores colheram dados sobre as aves dessa ilha. Norma Maciel desenvolveu por anos estudos com a avifauna local, merecendo destaque seu trabalho na Reserva Biológica da Praia do Sul (Maciel et al., 1984). Coelho et al. (1991), em seu trabalho com aves insulares do Estado do Rio de Janeiro, também apresentam dados sobre a Ilha Grande e pesquisadores da Universidade Estadual do Rio de Janeiro têm realizado estudos de ecologia e história natural de várias aves locais (p. ex. Alves & Duarte, 1996; Van Sluys et al., 1998; Almeida et al., 2000; Alves et al., 2001). Em outras ilhas da região, dados inéditos da avifauna terrestre foram apresentados no Plano de Manejo da Estação Ecológica dos Tamoios (Fapur/UFRRJ, Eletronuclear & Ibama, 2001).
Informações esparsas sobre aves da região também podem ser encontradas no livro “Ornitologia Brasileira” de Helmut Sick (1997) e em dezenas de artigos nacionais e internacionais, além de resumos de congressos brasileiros de zoologia e ornitologia (Pacheco et al., 1997).
Apesar da região sul-fluminense dispor de considerável material bibliográfico sobre suas aves terrestres, muitas informações referem-se a localidades distantes da área de influência do empreendimento ou não especificam precisamente o local onde os registros foram obtidos. Dessa forma, a coleta de novos dados e a elaboração de uma listagem mais direcionada para o conhecimento atual da avifauna local tornaram-se necessários.
Os resultados encontrados são fruto da compilação de dados de bibliografia e de informações não publicadas (listas de aves não-publicadas e comunicação pessoal) bem como de dados obtidos durante duas excursões às Áreas de Influência Direta de Angra 3. Espécies marinhas ou aquelas registradas exclusivamente em ilhas não foram incluídas nessa listagem mas estão presentes no item 7.3.2.2. Optou-se também por incluir apenas registros de aves em áreas de baixada e floresta sub-montana (50m - 500 m de altitude), sensu Veloso et al. 1991, locais onde a influência da instalação da Usina deve ser mais imediatamente percebida.
Uma vez que o objetivo principal das atividades de campo foi registrar o maior número possível de espécies, optou-se por adotar um método simples que permitisse obter rapidamente informações sobre a riqueza específica local. A coleta de dados consistiu em registrar as espécies detectadas, tipo de vegetação e altitude à medida que as transecções (trilhas e estradas previamente existentes) iam sendo percorridas. Os locais de amostragem foram selecionados a partir de informações de moradores locais e consulta a fotos aéreas, mapas e cartas topográficas do IBGE e Exército Brasileiro.
Priorizou-se a seleção de transecções cortando matas de encosta (Figura 66), já que as baixadas da região haviam sido bem investigadas em estudos anteriores.
As aves foram identificadas através da observação com binóculos ZEISS 8 x 30 e 10 x 40 e/ou registro auditivo de suas vocalizações. O emprego de guias de campo, como Dunning (1987), Ridgely & Tudor (1989, 1994) e Souza (1998), foram de grande valia na identificação de espécies crípticas. Vozes desconhecidas foram gravadas com gravadores ITT Studio Recorder 65 e DAT SONY TCD-D8 usando respectivamente os microfones AKG D1000E e Seinnheiser ME-67. A reprodução dessas vozes (técnica de “play-back”) foi utilizada como recurso para atrair as aves até o alcance da visão e permitir a sua identificação. Registros sonoros desconhecidos foram posteriormente identificados a partir da consulta ao acervo do Arquivo Sonoro Prof. Elias Coelho (ASEC) da Universidade Federal do Rio de Janeiro.
A nomenclatura científica adotada, bem como a ordem sistemática das famílias e espécies, seguem Sick (1997).
A caracterização das espécies quanto à sua distribuição altitudinal foi principalmente baseada no trabalho de Scott & Brooke (1985). Outras fontes bibliográficas foram consultadas para corroborar e/ou complementar os dados (Collar et al., 1992; Sick, 1997 e Bauer, 1999).
A situação de conservação das espécies foi estabelecida segundo os autores Collar et al. (1992) e Bergallo et al. (2000), reunidas em dois grupos: ameaçadas de extinção (Provavelmente Extinta=PEx, Em perigo=EP, Ameaçada=Am, Vulnerável=Vu, SD=Situação Desconhecida) e quase ameaçadas de extinção (QA). Quando houve discordância entre as classificações dos autores, foi atribuído o grau mais elevado em termos de ameaça.
Figura 66 - Locais de amostragem de aves terrestres 
Ao todo, foram registradas 331 espécies nas baixadas e matas sub-montanas (Anexo 11). Dentre estas, 16 espécies são consideradas ameaçadas de extinção globalmente e 26 espécies quase ameaçadas.
No estado do Rio de Janeiro, 20 espécies são consideradas ameaçadas de extinção e 22 espécies quase ameaçadas (Tabela 58). Em conjunto, isto é, ameaçadas globalmente e no estado do Rio de Janeiro, estas espécies representam 20,5% de toda a avifauna local. Dentre elas, merecem destaque o papa-formigas-de-cabeça-negra Formicivora erythronotos, espécie endêmica da Costa Verde, o não-pode-parar Phylloscartes paulistus, espécie só assinalada para a região nos últimos cinco anos (D.Buzzetti com. pess.) e o anambezinho Iodopleura pipra, cujos vários registros recentes tornam a área uma das mais importantes para sua conservação.
Além desses, as presenças do patinho-gigante Platyrinchus leucoryphus, do saí-de-pernas-pretas Dacnis nigripes e da saíra-sapucaia Tangara peruviana foram consideradas suficientes para que Angra dos Reis fosse tratada como uma das áreas-chave para a conservação das aves neotropicais (Wege & Long 1995).
Tabela 58 - Espécies ameaçadas de extinção, provavelmente ameaçadas de extinção e as espécies endêmicas de Mata Atlântica registradas na área sob a influência da Usina Angra 3
Clique aqui para visualizar a tabela 58
PEx: Provavelmente Extinta; EP: Em Perigo.; Am: Ameaçada; Vu: Vulnerável; QA: Quase Ameaçada; SD: Situação Desconhecida.
Outras 44 espécies foram registradas nas baixadas e florestas sub-montanas dos municípios de Angra dos Reis e Parati, fora da área de estudo (Tabela 59). Dada a semelhança da vegetação, topografia e condições climáticas de todos esses ambientes, sugere-se que a maioria dessas espécies muito provavelmente também deva ocorrer, ainda que ocasionalmente, na área de estudo. Apenas o gavião-pega-macaco Spizaetus ornatus, cujos últimos registros para esses municípios datam da década de 70 (Pacheco et al. 1997), deve estar atualmente extinto no estado do Rio de Janeiro (Bergallo et al., 2000). Esse gavião tem se tornado escasso ao longo de toda a sua distribuição (Sick, 1997) e provavelmente não suportaria as fortes pressões antrópicas a que estão atualmente sujeitas as terras baixas da Costa Verde.
Tabela 59 - Espécies registradas em baixadas e florestas sub-montanas dos municípios de Angra dos Reis e Parati, RJ, adjacentes à área de estudo
Clique aqui para visualizar a tabela 59
Terborgh (1977) menciona a dificuldade em se definir limites altitudinais de espécies da Floresta Atlântica do sudeste brasileiro. Nessa região, espécies tipicamente montícolas em sua distribuição setentrional estendem sua faixa de ocorrência até as planícies costeiras da Serra do Mar (Tabela 60). No litoral sul-fluminense e norte-paulista, espécies com distribução predominantemente serrana ocorrem na região de Angra dos Reis e Parati, praticamente ao nível do mar (Buzzetti, 1998). Esse fenômeno aumenta a riqueza e a diversidade da fauna nas baixadas e encostas baixas da Costa Verde.
Tabela 60 - Espécies tipicamente serranas em sua distribuição setentrional registradas na área de influência da Usina Angra 3
Clique aqui para visualizar a tabela 60
Entre as 18 espécies de aves de montanha residentes na área de estudo merecem destaque a choca-de-chapéu-vermelho Thamnophilus ruficapillus, o chupa-dente Conopophaga lineata, o ferreirinho-de-cara-canela Todirostrum plumbeiceps e a borralheira Mackenziaena severa.
Fora do período reprodutivo (abril a junho), a região recebe migrantes latitudinais, como a tesoura Tyrannus savana, o príncipe Pyrocephalus rubinus e guaracavas Elaenia spp. que deixam a pátria onde procriam no sul do país fugindo do inverno austral. Chegam também a região visitantes procedentes do norte durante o inverno setentrional (novembro a abril), como maçarico Charadrius semipalmatus, o piui-boreal Contopus borealis e a andorinha Hirundo pyrrhonota. A composição da avifauna local sofre ainda alterações durante os meses frios com a chegada de migrantes altitudinais. Espécies com distribuição predominantemente serrana, como a tesourinha Phibalura flavirostris, o pula-pula-assobiador Basileuterus leucoblepharus e a saíra-lagarta Tangara desmaresti, visitam as terras baixas da Costa Verde nos meses de julho a agosto, fugindo das baixas temperaturas das grandes altitudes.
O conhecimento da história natural dos animais pode ser de grande valia na elaboração de estratégias para sua conservação. Um dos aspectos da biologia das espécies mais relevante nesse sentido é a reprodução. Assim, por exemplo, o cronograma de atividades de manejo de determinada espécie ou comunidade deve levar em conta o respectivo período reprodutivo.
Mendonça (2001) apresenta dados sobre a reprodução das aves nas baixadas de Angra dos Reis e Parati em 1998 e 1999. As atividades reprodutivas da avifauna local se concentraram de meados de agosto a início de fevereiro (Figura 67). Como os dados não foram coletados sistematicamente, pequenas variações mensais podem ser reflexos de defeitos amostrais. Esse período coincide com o padrão geral observado para as aves do Hemisfério Sul, cujas atividades estão geralmente concentradas no final da estação seca e início da estação chuvosa (Skutch 1950, Pinto 1953, Snow 1976 e Sick 1997).
Figura 67 - Período de atividade reprodutiva (construção, incubação e cuidado com o filhote) da avifauna de baixada em Angra dos Reis e Parati, RJ, em 1998 e 1999.
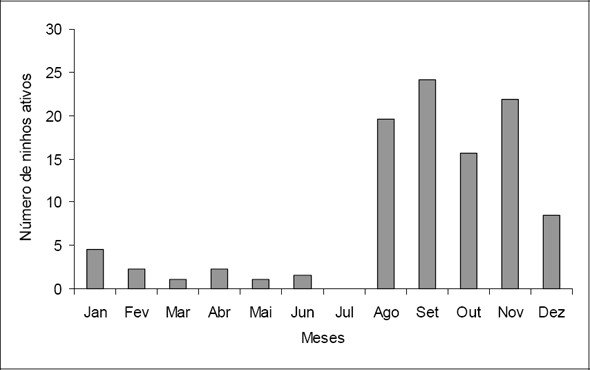
Exceções ao padrão reprodutivo da avifauna local foram observadas para insetívoros como o joão-botina Phacellodomus erythrophthalmus, as chocas Thamnophilus palliatus e Thamnophilus ruficapillus e a cambaxirra Troglodytes aedon (E. Mendonça obs. pess.). Essas variações podem ser resultado das atividades humanas na região, alterando o padrão normal de oferta de insetos no ambiente. Segundo Oniki & Willis (1982, 1983), aves granívoras e insetívoras com hábitos tipicamente sinantrópicos se beneficiam da disponibilidade constante de alimento produzida pela ação humana como a irrigação de plantas, podendo se reproduzir ao longo de todo o ano.
Um caso particular relacionado à conservação da avifauna na área de estudo diz respeito ao papa-formigas-de-cabeça-negra Formicivora erythronotos. Esta espécie permaneceu por mais de um século conhecida unicamente por algumas peles coletadas no século passado e depositadas em museus americanos e europeus (Pacheco, 1988). Após a sua redescoberta, a espécie permaneceu com registros esparsos de alguns poucos casais ao longo da Baía da Ribeira (Collar et. al.,1992; Tobias & Williams, 1996; Fernando Carvalho com. pess. 1999), o que levou a espécie a ser tratada como em situação crítica por Collar et al. (1992). Em 1997, pesquisadores do Laboratório de Ornitologia da UFRJ iniciaram uma procura sistemática do papa-formigas em todas as baixadas da região da Costa Verde, mapeando a população até então conhecida. Durante o estudo, constatou-se que o vale do Mambucaba e a baixada do Ariró, localidades inseridas na área de influência da Usina Angra 3, abrigam mais de 90% da população total da espécie (Mendonça & Gonzaga, 1999).
O vale do Mambucaba, que é a mais extensa planície da região, com cerca de 20 km2, é cortado pelo rio de mesmo nome, que serve como limite dos municípios de Parati e Angra dos Reis. O lado de Parati apresenta as maiores áreas contínuas de capoeira de baixada da região. Nesse local, F. erythronotos ainda pode ser encontrada com facilidade (densidade populacional de 156 casais/km2), sendo a mais importante área para a conservação da espécie (Mendonça & Gonzaga, 1999). O outro lado do vale do Mambucaba sofreu com a rápida urbanização após a implantação da CNAAA no início da década de 70 e teve grande parte de sua vegetação substituída por habitações e comércios. O interior do vale como um todo, abriga inúmeros sítios e fazendas, nos quais a vegetação original praticamente desapareceu. A banana, cultura predominante na região, representando cerca de 90 % da sua produção agrícola (MMA,1997), está extensamente distribuída em várias dessas propriedades. A desvalorização do produto nos últimos anos fez com que parte de seu cultivo fosse abandonado, dando origem a bananais “sujos” (entremeados de capoeiras), adequados à existência de F. erythronotos. Outra parte dos bananais tem cedido lugar às pastagens de gado e, principalmente, ao cultivo do palmito pupunha Euterpe sp., ambientes inóspitos à existência da espécie.
Na baixada do Ariró, a segunda maior planície da região, os cerca de 100 ha de terras baixas da Fazenda Ariró são cobertos em grande parte por um eucaliptal abandonado, cujo sub-bosque abriga F. erythronotos com densidade populacional de cerca de 89 casais/km2 (Mendonça e Gonzaga, 1999). O restante da baixada está quase totalmente inserido na Fazenda Pedra Branca, onde o eucaliptal, semelhante e outrora contínuo ao da Fazenda Ariró, está em franco declínio, dando lugar a pastagens de gado, que atualmente ocupam quase toda a sua planície. A área é atravessada por três linhas de transmissão de energia, as quais restringem o contato entre as duas sub-populações de F. erythronotos por uma faixa de cerca de 150 m de largura.
A destruição dos habitats, a caça, a introdução de predadores, competidores e doenças exóticas, têm sido as principais causas da extinção das aves. Particularmente no Rio de Janeiro, a destruição e fragmentação da Mata Atlântica são a principal ameaça para a maior parte da avifauna nativa (Alves et al., 2000). O tamanho dos fragmentos remanescentes pode não ser suficiente para abrigar espécies que exigem um espaço mais amplo para a sua sobrevivência. O isolamento de espécies em pequenas áreas separadas por grandes espaços de terras cultivadas ou pastagens impede-as de manter seu ciclo biológico ao longo do ano. Muitas espécies de tucanos, papagaios e cotingídeos, após a reprodução empreendem migrações locais geralmente em busca de certas frutas e sementes cuja ocorrência pode ser local e variável durante o ano (Sick, 1997).
Chiarello (2000) alerta que a curto prazo, a pressão de caça pode afetar mais intensamente as populações em fragmentos tropicais isolados do que por fatores biológicos “intrínsecos” como a competição e a predação. A caça e captura para cativeiro têm um efeito desproporcionalmente alto sobre alguns grupos, como os tinamídeos e cracídeos (caça), assim como sobre os psitacídeos e emberezídeos (cativeiro). O fato de a Mata Atlântica ter mais de 200 espécies endêmicas de aves (Stotz et al., 1996) aumenta o impacto global da devastação antrópica neste bioma, pondo em risco uma avifauna única (Alves et al. 2000).
Dessa forma, não somente o papa-formigas, mas também outras aves ameaçadas de extinção que ocupam ou ao menos visitam periodicamente as baixadas e encostas adjacentes, como o gavião-pomba Leucopternis lacernulata, a jacutinga Pipile jacutinga, o anambézinho Iodopleura pipra e a tesourinha Phibalura flavirostris, certamente também sofrem com o processo de descaracterização a que essas regiões estão atualmente sujeitas.
7.3.2.2. Caracterização da Avifauna Aquática - topo
O segmento de aves aquáticas é constituído por espécies que vivem em ambientes relacionados à água, tais como mares, lagoas, manguezais, brejos e banhados, podendo ser divididas em três grupos básicos: marinhas; dulcícolas; e mistas. As espécies marinhas apresentam hábitos essencialmente marinhos, vivendo e se reproduzindo em ambiente costeiro ou oceânico. As aves dulcícolas representam um grupo bem mais restrito, sendo constituído por espécies típicas de águas doces. As aves de ambientes aquáticos mistos formam um grupo bem mais numeroso, vivendo em ambientes aquáticos variados, podendo habitar tanto praias oceânicas quanto as margens de lagoas e lagos de água doce, manguezais, banhados, brejos ou mesmo pequenos riachos (Mallet-Rodrigues, 2001). Foram incluídos neste grupo toda a família Alcedinidae, apesar de Chloroceryle inda (martim-pescador-da-mata) ocorrer em matas de baixada e não em áreas alagadas como as outras espécies desta família.
No EIA da Unidade 2 da CNAAA - Angra 2 (NATRONTEC, 1999a) existem informações sobre o grupo de aves aquáticas, principalmente na área da Estação Ecológica de Tamoios. Dados sobre a ocorrência deste grupo para manguezais, restingas, mata atlântica e para a Ilha Grande encontram-se dispersos em meio a outros grupos zoológicos e foram obtidos através de consulta à literatura (Maciel et al., 1984, Maia & Oliveira, 1976, Rizzini et al. 1988, Sick, 1997), de algumas observações de campo e de entrevistas com mateiros da região.
No Projeto Básico Ambiental de Angra 2 (NATRONTEC, 1999b), que visou a complementação da caracterização da cobertura vegetal e da fauna associada, são citadas ocorrências de aves, dentre elas algumas de hábitos aquáticos. Estas listas foram baseadas em observações em campo, em pesquisas a coleções do Museu Nacional e em entrevistas com mateiros da região.
Na Estação Ecológica de Tamoios, o levantamento da avifauna aquática das 29 ilhas, ilhotes, lajes e rochedos, existentes nas baías da Ribeira e da Ilha Grande (área de influência direta), realizado em fevereiro de 1998, identificou 13 espécies de aves. Larus dominicanus (gaivotão), Sterna eurygnatha (trinta-réis-de-bico-amarelo), Sterna maxima (trinta-réis-real), Haematopus palliatus (ostreiro), Sula leucogaster (atobá-marrom) e Fregata magnificens (fragata) são as espécies de hábitos marinhos. As aves observadas que vivem em ambientes aquáticos mistos foram: Casmerodius albus (garça-branca-grande), Nyctanassa violacea (savacu-de-coroa), Nycticorax nycticorax (savacu), Ardea cocoi (socó-grande) e Ceryle torquata (martim-pescador-grande), além dosmigrantes do Hemisfério norte Actitis macularia (maçarico-pintado) e Arenaria interpres (vira-pedras).
Das ilhas que compõem a Estação Ecológica faltaram registros para Tucum de Fora, Palmas, Ganchos, Catimbaú e Laje Pedra Pelada. A Ilha Zatin, dentre as visitadas, teve importância para o grupo dos trinta-réis (Sterna spp) com um registro de cerca de 80 indivíduos de Sterna maxima (trinta-réis-real) e aproximadamente 500 indivíduos de Sterna eurygnatha (trinta-réis-de-bico-amarelo). As aves apresentavam a plumagem de descanso reprodutivo. Sterna eurygnatha é sensível a interferências humanas e suas populações necessitam proteção em seus poucos sítios reprodutivos conhecidos. É a espécie costeira mais ameaçada de extinção no Brasil (Antas,1991).
O Plano de Manejo da Estação Ecológica de Tamoios (FAPUR - UFRuralRJ - Eletronuclear - Ibama, 2001) fornece uma listagem de aves, resultado de consultas a dados publicados (Berla, 1944, Pacheco et al., 1997), a informações contidas no EIA de Angra 2 e a listas de aves não publicadas (Pedro Develey 1995, Dante Buzzetti em 1995 e Brandão em 1997).
Estudos na região citam a ocorrência de espécies aquáticas, como em Maciel et al. (1984) para a Reserva Biológica da Praia do Sul e em Coelho et al. (1991) para ilhas costeiras. A divulgação de dados inéditos para a região pode ser encontrada em Pacheco et al. (1997). Parte destas informações refere-se a material coletado por Berla nos anos 40 e por L. Travassos no período de 1910 a 1930. Mas recentemente, Buzzetti (2000) pesquisou diversas localidades nos municípios de Angra dos Reis e Parati.
Uma contribuição ao conhecimento de fauna e flora da Reserva Biológica da Praia do Sul, localizada na Ilha Grande, é encontrada em Maciel et al.(1984). Algumas espécies aquáticas foram registradas pelos autores como Sula leucogaster (atobá-marrom), Fregata magnificens (fragata), Larus dominicanus (gaivotão), Tigrisoma lineatum (socó-boi), Butorides striatus (socozinho), Egretta thula (garça-branca-pequena), Ceryle torquata (martim-pescador-grande), Chloroceryle americana (martim-pescador-pequeno) e Platalea ajaja (colhereiro). Esta última espécie, segundo Alves et al. (2000), consta da lista de aves provavelmente ameaçadas no Estado do Rio de Janeiro. Outras aves citadas para a localidade por Pacheco et al. (1997) são: Podiceps dominicus (mergulhão-pompom) e Ardea cocoi (garça-moura, maguari).
No estudo efetuado em ilhas costeiras do Estado do Rio de Janeiro (Coelho et al., 1991), existem informações sobre a ilha de Jorge Grego e a ilha Grande, ambas na região de Angra do Reis. Para Jorge Grego são citadas: Sula leucogaster; Fregata magnificens; Haematopus palliatus; Larus dominicanus; e Aramides cajanea. Em 1983, V.S.Alves, juntamente com Elias P. Coelho e Max Luiz L. Soneghet, registrou a reprodução de F.magnificens nessa ilha. Em ilha Grande, foram observadas: Sula leucogaster; Fregata magnificens; a saracura Laterallus viridis (siricora-mirim); Larus dominicanus; e Actitis macularia.
No já citado estudo de Pacheco et al. (1997) “Novos registros de aves para o Estado do Rio de Janeiro: Costa Verde”, que abrange os municípios de Parati e Angra dos Reis, incluindo as ilhas da Baía da Ilha Grande e a região da Reserva de Rio das Pedras em Mangaratiba, foram divulgados dados regionais inéditos. Dentre as espécies aquáticas, são citadas: Phalacrocorax brasilianus (biguá); Egretta caerulea (garça-azul); Syrigma sibilatrix (maria-faceira) como visitante ocasional; Pilherodius pileatus (garça-real); Anas bahamensis (marreca-toicinho); Netta peposaca (marrecão) como visitante ocasional; Amazonetta brasiliensis (marreca-pé-vermelho); Laterallus exilis (sanã-do-capim); Laterallus melanophaius (sanã-parda); Gallinula choropus (galinha-d’água); Porphyrula martinica (frango-d’água-azul); Charadrius semipalmatus (batuíra-de-bando); Calidris fuscicollis (maçarico-de-sobre-branco); Gallinago paraguaiae (narceja-comum); Rynchops niger (talha-mar-preto); e Chloroceryle amazona (martim-pescador-verde).
Buzzetti (2000) realizou um estudo sobre distribuição altitudinal de aves em Angra dos Reis e Parati, com enfoque em espécies terrestres. Entretanto, cita a ocorrência de algumas aves aquáticas, como Amaurolimnas concolor (Saracurinha-da-mata), Aramides mangle (saracura-da-praia) e Porzana albicollis (sanã-carijó).
O EIA de Angra 2 (NATRONTEC, 1999a) cita algumas espécies de aves aquáticas para o ecossistema manguezal, como na área do Hotel Pousada do Bosque, no km 143 da BR-101 e em Angra dos Reis, onde foram registradas, dentre outras: Rallus nigricans (saracura-sanã); Rallus longirostris (saracura-do-mangue); e Platalea ajaja (colhereiro).
É citada a presença de Jacana jacana (jaçanã), observada no manguezal da praia da Chácara, e de Platalea ajaja (= Ajaia ajaja), nos manguezais de Bracuí/Cansado (Natrontec, 1999b).
Para a área de influência indireta do empreendimento, há informações sobre Parati-Mirim, a 3 km de Parati. Em regiões alagadas de mata atlântica, ocorre o pato-do-mato (Cairina moschata), espécie muito perseguida por caçadores (Sick, 1997). Para as restingas, são citados Vanellus chilensis (quero-quero) e Dendrocygna viduata (marreca-irerê). Para manguezais do município de Parati, como Jabaquara, de Jorge Nunes, da Ponta da Navalha, do Saco Mamanguá e de Cairuçu, ocorrem os migrantes: Calidris alba (maçarico-branco); Tringa solitaria (maçarico-solitário); e Tringa flavipes (maçarico-de-perna-amarela), dentre outros registros (NATRONTEC, 1999a). Por fim, é citada uma listagem de aves para as áreas de influência direta e indireta do empreendimento, onde há o registro da saracura-do-mato (Aramides saracura) e do maçariquinho(Calidris minutilla), este visitante do hemisfério norte (NATRONTEC, 1999b).
As regiões pesquisadas em relação às espécies aquáticas foram principalmente as 29 ilhas, ilhotes e rochedos que compõem a Estação Ecológica de Tamoios e outras 37 ilhas e lajes, algumas das quais propícias à nidificação e/ou descanso de aves marinhas. Além destas áreas, foram feitas algumas observações em trechos de manguezais em Bracuí (Saco do Bracuí e Cansado) e Ariró (próximo à Ilha Comprida), onde foram registradas diversas espécies de aves aquáticas, que buscam neste ecossistema alimento, abrigo e locais para reprodução. As áreas amostradas estão apresentadas nas quatrto figuras a seguir.
Os trabalhos de campo foram realizados em três excursões. A região foi percorrida em um barco de 15 m, mantendo-se uma velocidade média 12 km/h. Os manguezais foram percorridos em bote inflável. Os pontos de embarque e desembarque foram o Porto de Bracuí e praia de Tarituba.
Figura 68 - Vista geral da região de Angra dos Reis e Parati, com as sub-regiões de amostragem (1a, 1b e 1c).
Figura 69 - Detalhe da sub-região 1a com as áreas amostradas
Figura 70 - Detalhe da sub-região 1b com as áreas amostradas
Figura 71 - Detalhe da sub-região 1c com as áreas amostradas
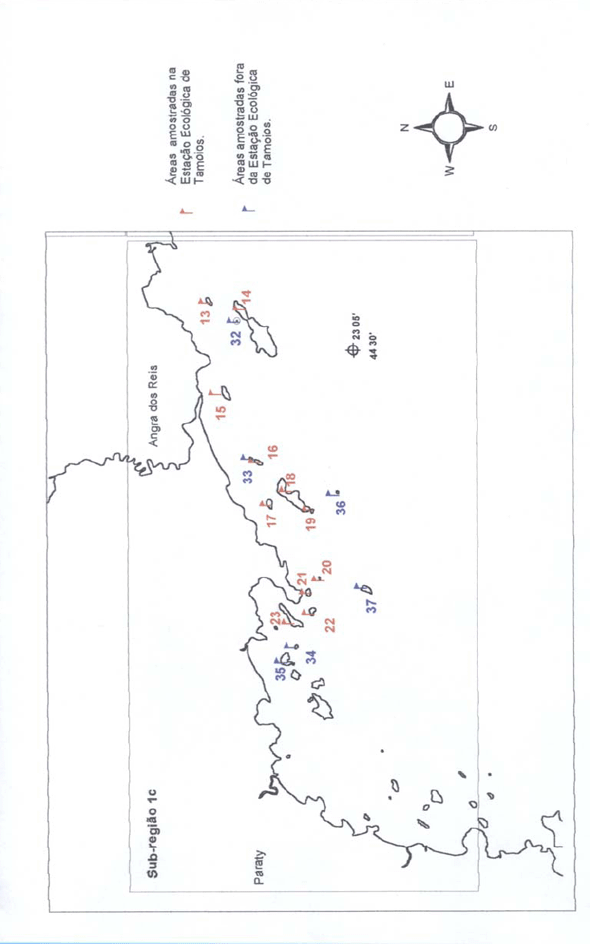
OBS.: Legenda com os nomes das ilhas, ilhotes, lajes, parcéis e rochedos amostrados na Estação Ecológica dos Tamoios
| Em vermelho nos mapas |
Em azul nos mapas |
| 1 - Ilha Tucum de Dentro |
1 - Ilha do Jorge |
| 2 - Parcel Pingo d’água |
2 - Ilha do Pasto |
| 3 - Ilha Tucum |
3 - Ilha Cunhambebe Grande |
| 4 - Ilha dos Búzios |
4 - Ilha do Sabacuzinho |
| 5 - Ilha dos Búzios Pequena |
5 - Ilhas Palmeiras |
| 6 - Ilha das Cobras |
6 - Ilha Itanhangá |
| 7 - Ilha Zatin |
7 - Ilha do Pinto |
| 8 - Ilha Imboacica |
8 - Ilha Paquetá |
| 9 - Ilha Queimada Grande |
9 - Ilhas Catitas |
| 10 - Ilha Queimada Pequena |
10 - Parcel do Pitanga |
| 11 - Ilha Araçatiba de Fora |
11 - Ilha Comprida |
| 12 - Ilha de Sabacu |
12 - Ilha do Maná |
| 13 - Ilha Samambaia |
13 - Lage do Aleijado |
| 14 - Ilha Sandri |
14 - Ilha do Aleijado |
| 15 - Ilha Algodão |
15 - Ilhas Cajeiras |
| 16 - Rochedos de São Pedro |
16 - Ilha do Major |
| 17 - Ilha Araraquarinha |
17- Ilha dos Coqueiros |
| 18 - Ilha Araraquara |
18 - Ilha do Cabrito |
| 19 - Ilha Jurubaíba |
19 - Laje Branca (próx. à Angra dos Reis) |
| 20 - Laje do Cesto |
20 - Ilha do Brandão |
| 21 - Ilhote Pequeno |
21 - Ilha do Pau a Pino |
| 22 - Ilhote Grande |
22 - Ilha da Josefa |
| 23 - Ilha Comprida |
23 - Lage de Búzios Norte |
| 24 - Ilha Araçatiba de Dentro |
24 - Ilha dos Papagaios |
| Obs. Ilhas Palmas, Ganchos, Catimbau e |
25 - Lage do Coronel |
| Laje Pedra Pelada - não amostradas |
26 - Ilha dos Porcos Grande |
| |
27 - Ilhas de São João |
| |
28 - Ilhas Botinas |
| |
29 - Ilha do Almeida |
| |
30 - Ilha da Piedade |
| |
31 - Ilha da Gipóia |
| |
32 - Ilha Mingu |
| |
33 - Rochedinho |
| |
34 - Ilha do Sururu |
| |
35 - Ilha Pelada Grande |
| |
36 - Laje Branca (próx. à Mambucaba) |
| |
37 - Ilha Araçatiba |
Em algumas ilhas houve desembarque como nos Rochedos de São Pedro, na Laje Branca (perto de Angra dos Reis), na Laje do Aleijado, na Ilha Zatin e na Ilha Pelada Grande. Entretanto, as observações, em geral, foram realizadas bordeando-se as ilhas.
As excursões para cada ilha variaram de uma a três visitas. As ilhas nas quais registraram-se concentrações de aves no primeiro dia, foram vistoriados nos dois dias subseqüentes; algumas, apesar de não apresentarem registros de aves, eram vistoriadas por estarem na rota do percurso.
Além de identificar as espécies, foram realizadas contagens diretas dos indivíduos, para possibilitar a identificação das áreas de relevância para o grupo pesquisado.
Os dados de campo foram gravados em fita com microgravadores (Olympus S700) e posteriormente transcritos. Para os registros fotográficos utilizou-se uma câmera fotográfica Olympus OM10 com objetivas de 50 mm e de 70 a 150 mm, grande angular de 28 mm e filmes fotográficos. Para marcação do trajeto e das coordenadas geográficas dos pontos amostrados foi utilizado um GPS Garmin 12.
Em laboratório, foram transcritas as informações que haviam sido gravadas sobre percursos, ilhas e manguezais pesquisados bem como dados quantitativos e qualitativos obtidos em cada área.
Os registros de campo, somados aos dados secundários já existentes para a região, considerando-se as áreas de influência do empreendimento, totalizam 56 espécies de aves aquáticas, conforme pode ser verificado na Tabela 61. A nomenclatura adotada está de acordo com Sick (1997).
Tabela 61 - Registros de espécies de aves aquáticas para a região de Angra dos Reis e Parati.
Clique aqui para visualizar a tabela 61
A Ordem Charadriiformes (representada principalmente pelos maçaricos migrantes e residentes) foi a mais diversificada, com 33,9 % do total das espécies registradas (56), seguida pelas Ordens Gruiformes (saracuras e frangos-d´água) com 21,4 % e Ciconiiformes (garças, socós e afins) com 19,6 %.
Os trabalhos de campo nas ilhas da Estação Ecológica de Tamoios, em outras 37 ilhas e lajes da região e nos manguezais do Saco do Bracuí, Cansado e Ariró listaram 24 espécies de aves aquáticas (Tabela 62), incluindo três novas ocorrências para a região: Pluvialis squatarola, Charadrius collaris e Chloroceryle aenea Conforme apresentado na Tabela 62, os dados revelam que a Ordem Charadriiformes, continua a ser a mais diversificada, com 12 espécies sendo sete migratórias representadas por batuíras e maçaricos e por espécies do gênero Sterna. As famílias Sulidae, Fregatidae, Haematopodidae e Laridae são representadas por espécies de hábitos marinhos.
Tabela 62 - Espécies de Aves aquáticas observadas nas áreas pesquisadas (Ilhas da Estação Ecológica de Tamoios, outras ilhas da região e nos manguezais de Bracuí (Saco do Bracuí e Cansado e Ariró).
Clique aqui para visualizar a tabela 62
Na Estação Ecológica de Tamoios foram observadas 15 espécies de aves aquáticas, sendo que ula leucogaster (atobá-marrom) e Fregata magnificens (fragata) foram mais freqüentemente observadas apenas sobrevoando as ilhas. A ocorrência destas aves em cada ilha, ilhote e rochedo que compõe esta Unidade de Conservação pode ser vista na Tabela 63.
Tabela 63 - Espécies de aves aquáticas observadas nas Ilhas da Estação Ecológica dos Tamoios.
Clique aqui para visualizar a tabela 63
De todas as áreas percorridas, Rochedos de São Pedro apresentou o maior número de espécies registradas seguida das ilhas Sabacu e Zatin. É importante ressaltar que todas as ilhas em questão são predominantemente rochosas. Geralmente áreas com pouca vegetação e de difícil acesso são locais favoráveis à reprodução e pouso de aves marinhas.
As aves marinhas registradas foram Sula leucogaster (atobá-marrom), Fregata magnificens (fragata), Haematopus palliatus (ostreiro), Larus dominicanus (gaivotão), Sterna hirundinacea (trinta-réis-de-bico-vermelho), Sterna eurygnatha (trinta-réis-de-bico-amarelo) e Sterna maxima (trinta-réis-real). As aves aquáticas foram: Ardea cocoi (socó-grande), Casmerodius albus (garça-branca-grande), Egretta thula (garça-branca-pequena), Vanellus chilensis (quero-quero), Charadrius semipalmatus (batuíra-de-bando), Arenaria interpres (vira-pedras), Actitis macularia (maçarico-pintado) e Ceryle torquata (martim-pescador-grande).
Embora sejam integrantes da Estação Ecológica de Tamoios, as ilhas Palmas, Catimbau e Ganchos não foram visitadas devido a considerável distância que as separa das outras ilhas estudadas. A Laje Pedra Pelada e a laje entre a Ilha de Búzios e a Ilha das Cobras não foram localizadas; talvez estivessem submersas durante o período.
Tabela 64 - Numero de indivíduos por espécies observados nas Ilhas da Estação Ecológica dos Tamoios.
Clique aqui para visualizar a tabela 64
Houve o registro de quatro espécies de maçaricos das quais três são migratórias, provenientes de regiões da América do Norte. Estas espécies, entretanto estiveram representadas por indivíduos isolados e principalmente nos Rochedos de São Pedro. As grandes concentrações destes migrantes em nosso país ocorrem durante a primavera e o verão.
Nos Rochedos de São Pedro foram observadas oito espécies de aves aquáticas, maior diversidade específica dentre os locais visitados; a espécie mais abundante foi o gaivotão Larus dominicanus (190 indivíduos)seguida pelas três espécies de trinta-réis.
Os trinta-réis, que foram observados em grandes concentrações (cerca de 500 indivíduos de S. eurygnatha e 80 S. máxima) na Ilha Zatin em fevereiro de 1998 (NATRONTEC, 1999a), estiveram ausentes desta ilha no período em que se realizaram as excursões deste trabalho, em maio de 2002. Neste período, a quantidade de indivíduos das três espécies de Sterna foi muito inferior, sendo registrados 71 indivíduos de Sterna máxima, 34 de S. eurygnatha e 21 de S. hirundinacea nos Rochedos de São Pedro.
O gaivotão (Larus dominicanus) foi observado em 12 das 24 ilhas visitadas, em geral em números pequenos (de um a 16 indivíduos), exceto nos Rochedos de São Pedro onde foram registrados 190 indivíduos. Em todas as observações havia indivíduos adultos e jovens. Esta espécie de gaivota é bastante comum e associada a áreas de ressurgência (Antas, 1991). Há registros reprodutivos no Rio de Janeiro na região de Cabo Frio (Sick, 1984).
As excursões para outras ilhas da região de Angra dos Reis objetivaram reunir mais informações sobre o grupo das aves aquáticas. Assim, foram pesquisadas 37 ilhas onde foram identificadas 19 espécies de aves aquáticas (Tabela 65).
Tabela 65 - Espécies de aves aquáticas observadas em outras ilhas da região
Clique aqui para visualizar a tabela 65
Assim como o observado nas ilhas da Estação Ecológica de Tamoios, S. leucogaster e F. magnificens, além de P. brasilianus encontravam-se geralmente sobrevoando a região.
As espécies de hábitos marinhos registradas foram Sula leucogaster (atobá-marrom), Haematopus palliatus (ostreiro), Larus dominicanus (gaivotão), Sterna hirundinacea (trinta-réis-de-bico-vermelho), Sterna eurygnatha (trinta-réis-de-bico-amarelo), Sterna maxima (trinta-réis-real). As aves de hábitos aquáticos foram: Ardea cocoi (socó-grande), Casmerodius albus (garça-branca-grande), Egretta thula (garça-branca-pequena), Egretta caerulae (garça-azul), Vanellus chilensis (quero-quero), Pluvialis squatarola (batuiruçú-de-axila-preta), Charadrius semipalmatus (batuíra-de-bando), Charadrius collaris (batuíra-de-coleira), Arenaria interpres (vira-pedras), Actitis macularia (maçarico-pintado) e Ceryle torquata (martim-pescador-grande).
Os maçaricos Pluvialis squatarola (migratório) e Charadrius collaris (residente) são ocorrências novas para a região; o primeiro foi registrado na Laje do Aleijado e o segundo na Laje Branca (próxima à Angra dos Reis).
As ilhas que apresentaram o maior número de espécies, considerando-se apenas as aves que estavam pousadas, foram a Laje do Aleijado e a Laje Branca - próxima à Angra dos Reis (ambas com nove espécies) e o Rochedinho - bem próximo aos Rochedos de São Pedro (com seis espécies). Assim como observado anteriormente, estes locais também são predominantemente rochosos e de difícil acesso.
A Tabela 66 mostra o número de indivíduos observados nas ilhas. Os trinta-réis - S.hirundinacea, S. eurygnatha e principalmente S.máxima foram aves comuns nas três áreas de maior diversidade específica. Na Laje Branca (próxima à Angra dos Reis), foi registrada a maior concentração de S.maxima (119 indivíduos), seguida da Laje do Aleijado (77) e do Parcel do Pitanga (28).
Os maçaricos das espécies C.semipalmatus, A. interpres, P.squatarola, A. macularia e C.collaris (este residente no Brasil)foram registrados principalmente na Laje do Aleijado, embora em baixo número.
Tabela 66 - Número de indivíduos por espécies observados em outras ilhas da região de Angra dos Reis
Clique aqui para visualizar a tabela 66
Não houve muitas observações em áreas de manguezais devido ao pouco tempo de campo e as condições adversas de tempo e visibilidade. Foram visitados trechos de manguezais em Bracuí (Saco de Bracuí e Cansado) e em Ariró (próximo à Ilha Comprida). A Tabela 67 apresenta as espécies registradas.
Tabela 67 - Aves aquáticas registradas nos manguezais do Ariró, Cansado e Saco do Bracuí
Clique aqui para visualizar a tabela 67
Os manguezais do Saco do Bracuí e Cansado apresentaram maior número de espécies registradas. Algumas utilizam este ecossistema como área de nidificação, repouso e alimentação. Para outras, como as aves migratórias, os manguezais funcionam como uma importante área, rica em alimento e onde também encontram abrigo.
Foram registradas 19 espécies de aves aquáticas para os manguezais estudados, sendo que seis possuem hábitos marinhos. Sula leucogaster e F.magnificens, não utilizavam propriamente os manguezais mas sobrevoavam as regiões; os trinta-réis (Sterna spp) aproveitavam as águas rasas para pescar.Um bando com mais de 100 indivíduos de S. máxima foi registrado no Saco do Bracuí.
A Ordem dos Ciconiiformes (garças e socós) esteve representada por seis espécies, muito comuns em áreas de manguezais, particularmente Egretta caerulae e N.violacea, esta última restrita ao litoral onde nidifica sobre árvores de mangue.
Apenas duas espécies de maçaricos migratórios foram registradas (A. macularia e A. interpres) o que certamente não reflete a realidade, pois estas aves são bastante comuns nestes ambientes no período de agosto/setembro a março/abril.
A ocorrência de Chloroceryle aenea (martim-pescador-anão) é nova para a região. Esta espécie habita margens de cursos d’água com densa vegetação, onde facilmente passa desapercebida (Sick, 1997).
As informações obtidas nas ilhas pesquisadas mostram a relevância de algumas destas áreas para aves do grupo dos trinta-réis (Sterna spp). Estas áreas foram os Rochedos de São Pedro, a Laje do Aleijado, a Laje Branca (próxima à Angra dos Reis), o Rochedinho e Parcel do Pitanga. Nas espécies de Sterna, o tamanho das populações nidificantes e os locais escolhidos para reprodução podem variar de ano para ano (Sick, 1997). Por não procriarem regularmente em determinadas áreas, as pesquisas sobre sua biologia reprodutiva são prejudicadas e muitas vezes esta característica torna difícil a preservação de suas áreas de concentração e reprodução.
No Brasil, as espécies de trinta-réis são comuns entre os meses de abril e setembro, freqüentando ilhas rochosas e sobrevoando regiões litorâneas em busca de alimento, que é constituído por pequenos peixes, seus alevinos e também crustáceos que capturam através do mergulho.
Os Trinta-réis (Sterna spp)
As três espécies de trinta-réis mais comuns no Rio de Janeiro, são freqüentemente observadas formando grandes bandos mistos em rochedos e ilhas. Nidificam em colônias e, algumas espécies, como S.hirundinacea e S.eurygnatha em simpatria. Surgem em nosso litoral por volta do mês de abril. Procriam no inverno e abandonam seus sítios reprodutivos por volta de setembro. No Rio de Janeiro S. eurygnatha e S. hirundinacea nidificam, por exemplo, na Baía de Guanabara (Maciel 1987, Maciel & Teixeira, 1984, Coelho et al., 1987). Até o momento não são conhecidos sítios reprodutivos do trinta-réis-real (S. máxima) no Estado do Rio de Janeiro.
Alguns predadores naturais de ovos e filhotes são o gaivotão (L. dominicanus), o urubu-comum (Coragyps atratus) e o gavião-caracará (Polyborus plancus). Além disso, interferências humanas são desastrosas e sempre causam o abandono do sítio e o comprometimento do ciclo reprodutivo.
Sterna maxima (trinta-réis-real) foi a espécie observada em maior número na região de Angra dos Reis. É um visitante regular proveniente de regiões setentrionais. Sabe-se muito pouco à respeito desta espécie. Escalante (1985) já comentava a falta de informações sobre o status dessa ave ao longo de mais ou menos 4000 km (da Guiana francesa até o Rio de Janeiro). Em 1984 as populações de Sterna maxima foram consideradas em perigo por Escalante (1984) pelo fácil acesso a suas áreas de nidificação, repouso e invernada, além da possibilidade de decréscimo no item que compõe sua dieta (peixe).
Há registros de indivíduos com plumagem de imaturo na Baía de Guanabara (obs.pess.) o que sugere a existência de um sítio reprodutivo relativamente próximo e ainda desconhecido. A literatura cita também o registro de aves em plumagem reprodutiva no Rio Grande do Sul em abril (Antas, 1991) e julho e setembro (Belton, 1984).
Recentemente foi descoberta a reprodução desta espécie no Brasil (Olmos et al.,1995), no litoral de São Paulo, mais especificamente na Laje de Santos.
Sterna eurygnatha (trinta-réis-de-bico-amarelo) é uma subespécie de Sterna sandvicensis, que possui ampla distribuição na América do Norte e na Europa (Sick, 1997). Na costa brasileira há registros de nidificação como na Baía de Guanabara (Maciel & Teixeira, 1984, Coelho et al., 1987), na Ilha dos Papagaios em Macaé (Sick & Leão, 1965) e principalmente nas ilhas costeiras do Espírito Santo (Moure et al., 1985, 1986; Nascimento et al., 1992; Efe & Musso, 1994; 1996, Efe et al., 2000). A época de nidificação desta espécie no Espírito Santo, segundo dados de Efe & Musso (1996), é entre maio e setembro. Foi considerada por Antas (1991) a espécie costeira brasileira mais ameaçada, quando o autor registrou que as colônias do Espírito Santo vinham sofrendo sérios impactos com a retirada de ovos por parte dos pescadores.
Sterna hirundinacea (trinta-réis-de-bico-vemelho) distribui-se da Terra do Fogo à Bahia. Nidifica na Baía de Guanabara (Maciel & Teixeira, 1984, Coelho et al. 1987), em ilhas costeiras do Espírito Santo (Moure et al., 1985, 1986 Efe & Musso, 1996), dentre outras regiões.
Migração
Aves limícolas (batuíras e maçaricos) migratórias com registro para a região são: Pluvialis squatarola (batuiruçú-de-axila-preta), Charadrius semipalmatus (batuíra-de-bando) ambos da Família Charadriidae, Arenaria interpres (vira-pedras), Tringa solitaria (maçarico-solitário), Tringa flavipes (maçarico-de-perna-amarela), Actitis macularia (maçarico-pintado), Calidris minutilla (maçariquinho), Calidris fuscicollis (maçarico-de-sobre-branco) e Calidris alba (maçarico-branco) daFamília Scolopacidae.
Estas aves, provenientes do hemisfério norte, chegam ao Brasil em agosto/setembro retornando em março/abril, para reproduzirem-se em seus países de origem (Sick, 1997). Permanecem, assim, em nosso território a maior parte do ano. Nos meses de verão, as batuíras e maçaricos migrantes costumam ser mais freqüentes e abundantes do que as próprias espécies residentes que normalmente são mais solitárias, formando grupos menores. Migram tanto durante o dia quanto de noite, percorrendo milhares de quilômetros, desde suas áreas de reprodução no norte da América do Norte.
Nesse estudo foram registradas apenas quatro das nove espécies que ocorrem na região; o período em que foi efetuado o estudo não era propício à observação destes migrantes.
As outras aves migratórias registradas foram os trinta-réis (Sterna spp). As três espécies mais comuns no litoral do Rio de Janeiro são S.hirundinaea, S.eurygnatha e S.maxima. No inverno, populações de S.hirundinaea e S.eurygnatha chegam ao Rio de Janeiro provenientes de regiões meridionais.
Hábitos alimentares
Dentre as 24 espécies observadas em campo, 16 são piscívoras e representadas pelas famílias Sulidae (uma espécie), Phalacrocoracidae (uma espécie), Fregatidae (uma espécie), Ardeidae (seis espécies), Laridae (quatro espécies) e Alcedinidae (duas espécies). Estas aves, no entanto, complementam a dieta consumindo itens como lulas, crustáceos, ovos e filhotes de outras aves marinhas, insetos aquáticos, anfíbios e répteis.
Os maçaricos identificados (quatro espécies migratórias e duas residentes) consomem invertebrados marinhos como poliquetas, crustáceos, moluscos, insetos aquáticos e suas larvas; algumas espécies também consomem insetos terrestres. Na periferia das ilhas estas aves costumam capturar a baratinha-da-praia (Lygia sp).
Aves aquáticas, de modo geral, são espécies indicadoras da qualidade ambiental das águas e dos ecossistemas associados; dependem, para sua sobrevivência, de recursos alimentares encontrados exclusivamente no meio aquático.
Espécies ameaçadas
Não há registros de aves aquáticas ameaçadas de extinção para a região. Por outro lado, ocorrem três espécies consideradas provavelmente ameaçadas no Estado do Rio de Janeiro (Alves et al., 2000) e outras três cujas informações atuais não permitem uma avaliação de seu status. Seria importante a realização de estudos populacionais sobre estas espécies.
As espécies provavelmente ameaçadas são Pilherodius pileatus (garça-real) - registros de Buzzetti (2000) e Pacheco et al. (1997), Platalea ajaja (colhereiro) - registros de Maciel et al. (1984) e NATRONTEC (1999a) e Rynchops niger (talha-mar) - registro de Pacheco et al. (1997). Pilherodius pileatus habita rios e lagos orlados de mata, é solitária, não sendo comum em lugar nenhum. Platalea ajaja ocorre em grande parte do Brasil, habitando manguezais e praias lamacentas. A destruição destes habitats torna a espécie mais escassa no litoral. Rynchops niger ocorre em rios e lagos do Brasil, migra no inverno, chegando ao litoral (Sick, 1997).
As outras três, cujo status é desconhecido são: Aramides mangle (saracura-do-mangue), registro de Buzzetti (2000), Chloroceryle inda (martim-pescador-da-mata), registro de Pacheco et al. (1997) e Chloroceryle aenea (martim-pescador-anão), observada no presente estudo.
Endemismos
Apenas uma espécie aquática listada para a área de estudo, Aramides mangle (saracura-do-mangue),é endemismo brasileiro e foi registrada por Buzzetti (2000). Esta saracura vive em praias lodosas com mangues e matas adjacentes segundo Sick (1997).
Fragilidades Ambientais
Áreas litorâneas vêm sendo destruídas a um ritmo intenso. A pressão sobre o nosso litoral é muito grande e inúmeros manguezais já foram destruídos. A região de Angra dos Reis não é diferente. Os manguezais foram ocupados por grandes empreendimentos como hotéis e marinas, que desmataram, aterraram e drenaram inúmeras áreas destes ambientes, principalmente no município de Angra dos Reis. Nos últimos 30 anos os manguezais foram reduzidos a 40% da área original e os existentes na área de influência direta do empreendimento representam 8,12% da área total do litoral sul do Estado do Rio de Janeiro e, de um modo geral, estão descaracterizados quanto ao seu estado nativo (NATRONTEC, 1999b).
Os ecossistemas insulares também sofrem pressões diversas; apesar de distantes e de acesso em geral difícil, recebem o impacto de atividades de pescas em seus arredores, perturbações em suas colônias de aves marinhas, uso de seus ovos como alimento, utilização de filhotes destas aves como iscas vivas, introdução de fauna e flora exótica, construções de moradias, desmatamentos etc
Aves terrestres observadasnas ilhas
As 43 espécies de aves de hábitos terrestres registradas nas ilhas estudadas estão listadas no Anexo 12. Quatro foram identificadas em nível de gênero.
Estas observações foram realizadas nas áreas de estudo e nas proximidades dos locais de embarque e desembarque (Porto de Bracuí e Praia de Tarituba). A maior diversidade de espécies foi (Ordem Passeriformes) foi a mais representativa, com maior diversidade de espécies nas famílias Emberizidae e Tyrannidae.
Nas ilhas pesquisadas, estas aves foram geralmente observadas pousadas em árvores, nas ilhas maiores como Gipóia, Comprida, Araraquara e Sandri, ou sobre arbustos, em ilhas menores como Botinas, São João, Samambaia e Algodão.
Nos manguezais, algumas aves terrestres buscam alimento (geralmente insetos), locais de pouso e reprodução. No Ariró foram encontrados ninhos vazios de Fluvicola nengeta (lavadeira-mascarada), Pitangus sulphuratus (bem-te-vi) e Cacicus haemorrous (guaxe). Um importante registro foi a vocalização de Procnias nudicollis (araponga) próximo a este manguezal, espécie provavelmente ameaçada de extinção no Estado do Rio de Janeiro (Alves et al., 2000).
Considerações sobre as aves aquáticas observadas
Sula leucogaster - atobá-marrom
Espécie tropical e subtropical que atinge o Paraná e Santa Catarina podendo chegar à Argentina. É uma ave de hábitos marinhos que nidifica principalmente em ilhas costeiras. Alimenta-se de peixes e lulas que captura através do mergulho. Adultos e filhotes foram freqüentemente observados sobrevoando e pescando próximos às ilhas, pousados em bóias sinalizadoras (nas proximidades da Laje do Coronel) ou acompanhando barcos de pesca. Apenas na Laje Branca (próxima a Mambucaba) foram vistos, nas duas excursões, adultos e imaturos pousados sobre arbustos, sugerindo ser um local de nidificação.
Phalacrocorax brasilianus - biguá
Ave aquática com ampla distribuição no Brasil, em lagoas, rios e outras áreas alagadas próximas da costa. Alimenta-se de peixes e também de camarões, que captura mergulhando em águas rasas. Registro de um indivíduo sobrevoando a Ilha do Almeida e outro no manguezal do Saco do Bracuí.
Fregata magnificens - fragata
Ave de hábitos marinhos com grande ocorrência na costa brasileira, nidificando em ilhas. Alimenta-se geralmente de peixes que captura na superfície da água ou rouba de outras aves marinhas, perseguindo-as em pleno vôo. Como os atobás, foi a espécie mais observada na região sobrevoando as ilhas e acompanhando barcos de pesca (indivíduos adultos e imaturos). Na Laje Branca (próxima à Mambucaba), foram vistos adultos com o bolsa gular vermelha e cerca de 12 indivíduos imaturos pousados em arbustos. Um dos adultos apresentava o comportamento de inflar a bolsa gular, característica de época reprodutiva. Há registros de nidificação na Ilha do Jorge Grego (obs.pess.).
Ardea cocoi - socó-grande
Garça de grande porte, que ocorre em todo o Brasil. Possui hábitos solitários e alimenta-se de peixes, caranguejos, moluscos, insetos aquáticos, e mesmo animais mortos. Foram observadas isoladamente na Ilha do Pau a Pino, pousada sobre uma bóia sinalizadora próxima às Ilhas Cajeiras e dois indivíduos na Ilha de Búzios.
Casmerodius albus - garça-branca-grande
Garça branca de grande porte, ocorre em todo o Brasil. Nidifica em ninhais, entre julho-agosto, em ilhas, brejos, manguezais e campos inundáveis, geralmente com outras espécies de garças e outras aves aquáticas. Efetua deslocamentos. Alimenta-se de peixes, insetos aquáticos, crustáceos, anfíbios etc. Poucos indivíduos pousados nas ilhas do Jorge, dos Porcos, Sandri, Comprida, Araçatiba de Dentro e do Sururu, além de presente no manguezais do Ariró, Cansado e Saco do Bracuí .
Egretta thula - garça-branca-pequena
Garça branca de pequeno porte, encontrada em todo o Brasil. Mais comum do que a espécie anterior, registros para as ilhas do Jorge, Tucum, Piedade, da Josefa, Sandri, do Major, Comprida, Araçatiba, Jurubaíba, Araraquara, Araçatiba de Dentro, Catitas e Ilhote Pequeno, em ruínas próximas ao Porto Bracuí e também nos manguezais do Ariró, Cansado e Saco do Bracuí.
Egretta caerulea - garça-azul
Garça cinza-azulada, que ocorre em todo o litoral brasileiro, sendo bem adaptada aos manguezais. Alimenta-se de crustáceos, pequenos peixes e insetos aquáticos. Com apenas um registro na Ilha do Jorge, mais comum nos manguezais do Ariró, Cansado e Saco do Bracuí.
Nyctanassa violacea - savacu-de-coroa
Espécie restrita ao litoral onde nidifica nos manguezais, ocorrendo no Brasil meridional até o norte do Rio Grande do Sul. Alimenta-se de pequenos peixes e insetos aquáticos. Vive nos manguezais empoleiradas. Foram observados alguns indivíduos nos manguezais do Ariró, onde houve o registro de quatro ninhos vazios, do Saco do Bracuí e Cansado.
Nycticorax nycticorax - savacu
Ocorre em quase todo o Brasil. É uma garça de hábitos noturnos e crepusculares. É comum empoleirar-se em árvores de manguezal. Alimenta-se de pequenos peixes. Registrada no manguezal de Cansado em Bracuí.
Aramides cajanea - saracura-três-potes
Saracura encontrada em todo o Brasil. É onívora, consumindo insetos, sementes, podendo predar pequenos vertebrados e ninhos de outras aves. Registro auditivo e visual no manguezal de Cansado.
Jacana jacana - jaçanã
Ave paludícola que ocorre em todo o Brasil, em brejo e lagoas, onde anda sobre vegetação aquática auxiliada pelos longos dedos e unhas. Alimenta-se de insetos, pequenos peixes, moluscos etc. Foi vista no manguezal do Saco do Bracuí.
Haematopus palliatus - ostreiro
Ave de hábitos marinhos que busca na beira-mar e em rochedos o alimento, que constitui-se de cracas e moluscos. Foram registrados indivíduos nas ilhas Zatim, Sabacu, Laje Branca - perto de Angra dos Reis (dois adultos e um jovem), Laje do Aleijado, Araraquara, Rochedos de São Pedro, Rochedinho, Laje Branca (próxima à Mambucaba) e Ilha Araraquarinha. Geralmente encontrados junto aos maçaricos, gaivotas e trinta-réis.
Vanellus chilensis - quero-quero
Maçarico residente de médio porte, encontrado em quase todo o Brasil, tanto em áreas costeiras como interiores. Alimenta-se de pequenos peixes, crustáceos, moluscos, insetos e outros artrópodes. Foram registrados indivíduos isolados nos Rochedos de São Pedro e Rochedinho.
Pluvialis squatarola - batuiruçu-de-axila-preta
Maçarico que ocorre em praias e em toda a costa atlântica do Brasil, chegando à Argentina, não penetrando no interior do continente. Ocorreu apenas um registro na Laje do Aleijado.
Charadrius semipalmatus - batuíra-de-bando
Maçarico que ocorre em praias lodosas e arenosas do litoral, em toda a costa brasileira até a Argentina. Foram registradas nas ilhas Zatim, Laje Branca (próxima à Mambucaba), Laje do Aleijado (máximo de 18 indivíduos alimentando-se), Rochedos de São Pedro e Sabacu. Sempre observado junto a outros maçaricos, gaivotas e trinta-réis.
Charadrius collaris - batuíra-de-coleira
Maçarico residente no Brasil. Apenas um registro na Laje Branca (próxima à Mambucaba).
Arenaria interpres - vira-pedras
Maçarico comumente observado na orla marítima rochosa onde captura moluscos e pequenos crustáceos. Registros de indivíduos na Laje Branca (próxima à Mambucaba), Laje do Aleijado, Rochedos de São Pedro, Rochedinho e Sabacu.
Actitis macularia - maçarico-pintado
Maçarico que ocorre na maior parte do Brasil, em margens lodosas e pedregosas de rios, freqüente em manguezais. Alguns registros em ilhas como Zatim, Laje Branca (próxima à Mambucaba) e Sabacu.
Larus dominicanus - gaivotão Gaivota comum no litoral brasileiro meridionalmente até o Rio de Janeiro. É onívora, sendo atraída por peixes mortos em praias; também predam ovos de outras aves marinhas. Nidifica, no inverno, em ilhas próximas ao continente. Adultos e jovens foram observados em rochedos e pousados nas ilhas. Grande número (190 aves) foi registrado nos Rochedos de São Pedro.
Sterna hirundinacea - trinta-réis-bico-vermelho
Trinta-réis que ocorre da Terra do Fogo à Bahia. Alimenta-se de peixes, alevinos e crustáceos que captura através do mergulho. Efetua migrações.
Sterna eurygnatha - trinta-réis-bico-amarelo
Trinta-réis que ocorre da Bahia ao Rio Grande do Sul. Alimenta-se de peixes, alevinos e crustáceos. Segundo a maioria dos autores recentes, é uma subespécie dotrinta-réis-de-bando (Sterna sandvicensis), amplamente distribuída na América do Norte e na Europa. Efetua migrações.
Sterna maxima - trinta-réis-real
Trinta-réis de grande porte, que ocorre no hemisfério sul onde reproduz-se até a Argentina. Alimenta-se de peixes e crustáceos. Efetua migrações.
Ceryle torquata - martim-pescador-grande
Ave aquática que ocorre em todo o Brasil, em beira de rios e lagos, em manguezais e próximo ao mar. Alimenta-se de pequenos peixes e também de insetos. Foram registrados isoladamente nas ilhas com vegetação mais alta como Queimada Grande, das Palmeiras, do Algodão, Cunhambebe Grande, de Búzios Pequena, Araçatiba de Fora e Araraquarinha e nos três manguezais pesquisados.
Chloroceryle aenea - martim-pescador-anão
Ave aquática com ocorrência em todo o Brasil. Alimenta-se de peixes e insetos. Registro para o Manguezal de Cansado.
Considerações sobre os registros
Foram registradas 56 espécies de aves aquáticas para Angra dos Reis e Parati; destas, 24 foram observadas nos trabalhos de campo realizados nas ilhas da Estação Ecológica, em outras ilhas da região e nos manguezais de Ariró, Cansado e Saco do Bracuí.
A Ordem mais diversificada foi a Charadriiformes com 12 espécies sendo sete migratórias representadas por batuíras e maçaricos e por espécies do gênero Sterna.
Na área da Estação Ecológica de Tamoios a maior diversidade de espécies aquáticas foi obtida nos Rochedos de São Pedro, seguido das ilhas Sabacu e Zatin. A espécie mais abundante foi o gaivotão Larus dominicanus, seguida pelas três espécies de trinta-réis (Sterna maxima, Sterna eurygnatha e S.hirundinacea).
Fora da Estação Ecológica, as ilhas com maior número de espécies foram: a Laje do Aleijado e a Laje Branca - próxima à Angra dos Reis; e o Rochedinho - bem próximo aos Rochedos de São Pedro.
Três espécies observadas (Pluvialis squatarola, Charadrius collaris e Chloroceryle aenea) constituem-se em novas ocorrências para a região.
Pilherodius pileatus (garça-real), Platalea ajaja (colhereiro) e Rynchops niger (talha-mar) são espécies provavelmente ameaçadas de extinção no Estado do Rio de Janeiro;
Aramides mangle (saracura-do-mangue), endemismo brasileiro, Chloroceryle inda (martim-pescador-da-mata) e Chloroceryle aenea (martim-pescador-anão) são espécies cujo status populacional encontra-se indefinido por falta de informações;
Maiores informações sobre os trinta-réis (Sterna spp) seriam obtidas, inclusive a possibilidade de registros inéditos de reprodução de S.maxima para o Estado do Rio de Janeiro, se os trabalhos de campo pudessem se estender ao período de junho-setembro.
Da mesma forma, a diversidade e a abundância dos maçaricos migratórios foram subestimadas, em decorrência da época do ano em que foram feitos os levantamentos de campo (outono-inverno).
7.3.3. Mastofauna - topo
O inventário de dados existentes sobre a mastofauna da área de influência de Angra 3 objetivou apresentar o estado de conhecimento atual da mastofauna daquela região, e está fundamentado em trabalhos recentes e na identificação de material depositado nas coleções do Museu Nacional e do Museu de Zoologia da Unicamp.
Este inventário subsidiou as atividades de campo desenvolvidas em 2002, quando se reuniu informações complementares para uma avaliação dos possíveis efeitos da implantação da usina sobre a composição das comunidades de pequenos mamíferos da região, bem como futuras proposições de medidas mitigatórias aos efeitos do funcionamento da usina no que diz respeito aos mamíferos especificamente.
Dada a impossibilidade de se efetuar coletas na ampla região ao longo de toda a área sob influência do projeto, optou-se por apresentar aqui um inventário baseado nos trabalhos que vêm sendo realizados nesta região e no material já depositado em coleções, para que as atividades de campo propriamente pudessem se concentrar em uma área mais restrita em torno do ponto de instalação da usina.
A coleção do Museu Nacional foi estudada com o intuito de se identificar os mamíferos da região da Baía da Ilha Grande e localidades adjacentes, em um raio de aproximadamente 20 km da área da usina, abrangendo, dessa forma, as áreas de influência direta. Um levantamento bibliográfico foi também realizado e parte das ocorrências de mamíferos relacionadas a seguir foram feitas a partir de informações constantes nestas publicações, tendo sido os nomes das espécies atualizados segundo Wilson & Reeder (1993) ou segundo revisões mais recentes.
7.3.3.1. Caracterização da Mastofauna - topo
As atividades de campo ocorreram em uma campanha de 10 dias, entre 18 e 28 de maio de 2002, com coletas diretas localizadas em três diferentes pontos amostrais: “Mata do Mamede” (Ponto Z4), “Torre de Microondas” (Ponto Z3a) e “Trilha da Telemar” (Z3b).
Coleta de pequenos mamíferos não voadores (marsupiais e roedores)
Para cada localidade de amostragem, foram estabelecidos sistemas de transecções isoladas, constituídas por armadilhas para coleta de animais vivos e de ratoeiras.
Nesta etapa foram utilizadas 44 armadilhas tipo “Sherman” dobráveis, 40 armadilhas de arame dobráveis e 50 ratoeiras japonesas de plástico dispostas no solo próximas a tocas e sobre árvores (Figura 72).
As armadilhas foram iscadas com aipim, aveia, banana, bacon e sardinhas. Fitas vermelhas numeradas foram usadas para marcação dos locais das armadilhas. Todas as armadilhas foram visitadas todas as manhãs para reiscagem e retirada dos animais capturados.
Indivíduos que não foram destinados à coleção foram devolvidos ao ponto de coleta original, depois de recolhidas informações que possibilitassem sua identificação inequívoca, bem como anotados os indícios externos que permitiram avaliar a idade e atividade reprodutiva.
Figura 72 - Tipos de armadilhas utilizadas em campo para coleta de pequenos Mamíferos

Coleta de quirópteros (morcegos) com redes de neblina
Para a captura de quirópteros foram utilizadas 4 redes de neblina (“mist-nets”) em cada um de 4 diferentes pontos georreferenciados, na “Mata do Mamede” e na “Trilha da Telemar”. Algumas redes foram dispostas próximas à inflorescências para a captura de morcegos nectarívoros e polinívoros e à árvores frutíferas na “Mata do Mamede” para captura de frugívoros. As redes ficaram montadas todos os dias de 17:00 às 21:00 horas.
Levantamento de indícios da presença de mamíferos (rastros, carcaças em decomposição ou partes do esqueleto, fezes)
Foram anotados, e quando possível, recolhidos ou fotografados, todos os indícios da presença de mamíferos. Espécimens encontrados mortos foram preservados em álcool para preparação do esqueleto.
Análise morfológica
No laboratório, os espécimens capturados foram fotografados para a composição de um arquivo de imagens dos animais da região e para dirimir possíveis dúvidas com relação à colocação da pelagem no caso dos animais preservados em meio líquido. Alguns dos espécimens coletados foram destinados à formação de uma coleção de referência. Para este fim, foram taxidermizados ou preparados em meio líquido no campo, seus esqueletos foram preparados posteriormente.
A identificação das espécies foi realizada com auxílio de chaves de identificação, revisões atualizadas e comparações direta com espécimens de regiões próximas já depositados no Museu Nacional. Os quirópteros coletados foram preparados em líquido e alguns crânios foram posteriormente retirados para confirmação da identificação. A coleção de referência será depositada na coleção de mamíferos do Museu Nacional como testemunho da identificação e da diversidade e no sentido de viabilizar análises da variabilidade morfológica visando a elaboração de hipóteses de diferenciação geográfica entre as amostras obtidas e as séries já disponíveis neste e em outros museus.
Análise Citogenética
Após serem coletados, alguns dos espécimens foram selecionados, mantidos em gaiolas criadeiras e transportados para o Laboratório de Mastozoologia da UFRJ. No laboratório foram feitas preparações cromossômicas de 13 indivíduos, sendo 7 roedores, 5 quirópteros e 1 marsupial. Para uma parte selecionada desses indivíduos coletados estão sendo preparados os cariótipos convencionais e da montagem para estimar o número cariotípico. As fases posteriores da análise dos cariótipos ainda encontram-se em processamento para a determinação do número diplóide (2n) e da morfologia cromossômica, através de bandamentos C, G, NOR.
A seguir, está relacionada a mastofauna documentada em acervos e em trabalhos realizados na região, em listas organizadas por localidades ou conjunto de localidades próximas. Para possibilitar comparações, as espécies foram relacionadas em duas listas, a primeira constando das localidades no continente, e a segunda relacionando as amostras obtidas até o momento na Ilha Grande. Foram constatados representantes de 9 ordens e aproximadamente 77 espécies de mamíferos silvestres na área de influência da Usina Angra 3. A Tabela 68 e a Tabela 69 listam as espécies nas diferentes localidades estudadas.
Tabela 68 - Espécies de mamíferos registradas em localidades continentais da área de influência de Angra 3.
Clique aqui para visualizar a tabela 68
Tabela 69 - Espécies de mamíferos da área de influência de Angra 3. Espécies registradas em localidades da Ilha Grande.
Clique aqui para visualizar a tabela 69
Na Figura 73 apresenta-se a distribuição potencial das espécies de marsupiais presentes na área de influência direta da usina, extrapolada através do georeferenciamento dos pontos de captura.
Figura 73 - Localidades de coleta de marsupiais (M)
Na Figura 74 apresenta-se a distribuição potencial das espécies de Chiroptera (morcegos) presentes na área de influência direta da usina, extrapolada através do georeferenciamento dos pontos de captura.
Figura 74 - Localidades de coleta de Chiroptera © (morcegos)
Na Figura 75 apresenta-se a distribuição potencial das espécies de roedores presentes na área de influência direta da usina, extrapolada através do georeferenciamento dos pontos de captura.
Figura 75 - Localidades de coleta de Roedores (R)
A seguir, apresenta-se lista preliminar georeferenciada das espécies de mamíferos coletadas (C) fotografadas (F) observadas (O), ou ainda registradas indiretamente por indícios da presença (I) - Tabela 70.
Tabela 70 - Vinte espécies de mamíferos registradas na região de influência da Usina Nuclear Angra 3 durante o período de 18 a 28 de maio de 2002.
Clique aqui para visualizar a tabela 70
Considerações sobre mamíferos observados
Ordem Didelphimorphia
Família Didelphidae
Didelphis aurita (gambá)
Somente um macho foi coletado no ponto Z3 (d) e cariotipado no laboratório. Dez outros espécimens foram capturados e liberados no ponto de coleta após identificação. Didephis aurita é um marsupial de hábitos noturnos e crepusculares, de dieta onívora, com destaque para insetos e frutos, e habitante de áreas de floresta atlântica, com grande tolerância a ambientes modificados pela ação antrópica. Sua relativa abundância nas áreas estudadas é congruente com o que vem sendo observado em outras áreas na Floresta Atlântica, e provavelmente é função escassez de espécies de predadores de maior porte, sendo que os maiores indivíduos desta espécie provavelmente encontram-se entre os predadores mais significativos em áreas de floresta sob moderado impacto antrópico.
Ordem Chiroptera
Família Phyllostomidae
Subfamília Stenodermatinae
Artibeus lituratus (morcego frugívoro)
Foram coletados seis indivíduos, sendo duas fêmeas em redes montadas na “Mata do Mamede” (trilha Z4) e na mata atrás do complexo nuclear (“Trilha Telemar”). Todos os espécimes foram considerados adultos de acordo com o graus de ossificação das falanges dos dígitos dos membros anteriores. Três indivíduos selecionados tiveram o crânio retirado para os procedimentos de identificação. Esta espécie tem sido abundantemente registrada em áreas de floresta atlântica, sendo muito comum em áreas de ocupação humana. Suas densidades populacionais em uma determinada área são reflexo da disponibilidade de recursos (Sazima et al., 1994). As espécies deste gênero desempenham um papel importante na dispersão de sementes de um variado número de espécies vegetais e na manutenção das florestas tropicais (Fleming, 1987; Marques-Aguiar, 1994).
Artibeus fimbriatus (morcego frugívoro)
Foram coletados dois machos em rede montada na “Mata do Mamede” (trilha Z4). Um indivíduo teve o crânio retirado para os procedimentos de identificação. Todos os indivíduos foram considerados adultos de acordo com o graus de ossificação das falanges dos dígitos dos membros anteriores. A biologia desta espécie é ainda pouco conhecida (Marquez-Aguiar, 1994). Assim como a espécie anterior, indivíduos desta espécie atuam como importantes dispersores de diversas espécies vegetais e são essenciais na manutenção das florestas tropicais (Fleming, 1987; Gardner, 1977).
Platyrrhinus lineatus (morcego frugívoro)
Coletadas duas fêmeas em redes montadas na mata atrás do complexo nuclear (“Trilha Telemar”). Os dois espécimes foram considerados adultos de acordo com o grau de ossificação das falanges dos membros anteriores. Os espécimes foram coletados em redes montadas próximas a figueiras (Ficus sp.) em fase de frutificação. Este morcego é considerado um importante dispersor de sementes. Tem sido observado na literatura a associação desta espécie de morcego com figueiras (Ficus sp.) (Gardner, 1977).
Subfamília Glossophaginae
Anoura caudifera (morcego nectarívoro)
Coletados quatro machos (um com testículos abdominais e três com testículos descendentes) em redes montadas na “Mata do Mamede” (trilha Z4) e na mata atrás do complexo nuclear (“Trilha Telemar”). Todos os espécimens foram considerados adultos de acordo com o grau de ossificação das falanges dos membros anteriores. Um espécimen teve o crânio retirado para identificação. Um espécimen coletado na mata atrás do complexo nuclear apresentou uma listra branca irregular na pelagem na região facial. Todos os espécimens foram coletados nas proximidades de bananeiras (Musa paradisiaca) em fase de floração. A. caudifera é um importante agente polinizador de diversas especies vegetais, inclusive as de interesse econômico. A utilização da bananeira como recurso por este morcego ja foi citada na literatura (Gardner, 1977).
Subfamília Lonchophyllina
Lonchophylla mordax (morcego nectarívoro)
Coletados dois machos (um com testículos abdominais e um com testículos descendentes) em redes montadas na mata atrás do complexo nuclear (“Trilha Telemar”). Os dois espécimens foram considerados adultos de acordo com o grau de ossificação das falanges dos membros anteriores. Um espécimen teve o crânio retirado para os procedimentos de identificação. Os espécimens foram coletados próximos a bananeiras em fase de floração. Pouco se conhece sobre a biologia desta espécie, entretanto tem sido observada sua associação com espécie vegetais de importância econômica (Nowak, 1993). O uso de bananeiras como recurso por esta espécie de morcego já foi observado em outras regiões ao longo de sua distribuição geográfica (Gardner, 1977).
Subfamília Sturnirinae
Sturnira lilium (morcego frugívoro)
Coletados três espécimens, sendo um macho e duas fêmeas em redes montadas na mata atrás do complexo nuclear (“Trilha Telemar”). Todos os espécimens foram considerados adultos de acordo com o graus de ossificação das falanges dos membros anteriores. Esta espécie tem sido observada associada com plantas pioneiras e plantas de interesse econômico (Gardner, 1977; Marinho-Filho, 1991). Esta espécie desempenha um papel importante na dispersão de sementes de um grande número de espécies vegetais Gardner, 1977; Marinho-Filho, 1991; Muller & Reis, 1992).
Subfamília Carolliinae
Carollia perspicillata (morcego frugívoro)
Coletados 14 indivíduos, sendo cinco machos e nove fêmeas foram coletados rede montada na “Trilha Z4” e na “Trilha Telemar”. Três indivíduos (um macho e duas fêmeas) foram considerados jovens por não apresentarem as falanges dos dígitos dos membros anteriores com ossificação incompleta. Os demais indivíduos foram considerados adultos. Todos os espécimens foram coletados em redes montadas nas proximidades de plantas em frutificação (Piper sp. e Solanum sp.). Esta espécie é uma das mais abundantes em florestas secundárias onde ocorre em associação com plantas pioneiras (famílias Piperaceae e Solanaceae) (Marinho-Filho, 1992; Cloutier & Thomas, 1992). A dispersão de sementes por esta espécie de morcego tem sido considerada essencial para a regeneração de áreas degradas ou com elevado impacto ambiental (Fleming, 1987).
Subfamília Desmodontinae
Desmodus rotundus (morcego hematófago)
Somente o indício da presença foi detectada durante uma visita para a seleção de pontos de coletas na fazenda Nova Grataú, localizada na margem esquerda do rio Grataú. Neste local, alguns indivíduos do rebanho de gado apresentavam marcas de mordeduras resultantes do ataque de morcegos hematófagos. Este fato foi confirmado pelos empregados da fazenda que relataram freqüentes ataques aos animais. Embora tenham sido montadas redes no local nenhum espécime foi capturado e não houve ataque nas noites em que a equipe esteve no local. Este morcego é considerado um dos maiores transmissores do vírus rábico e tem sido responsável por grandes perdas na indústria pecuária (Sazima, 1978).
Família Vespertilionidae
Myotis nigricans (morcego insetívoro)
Dois indivíduos, um macho e uma fêmea, foram coletados em redes montadas na mata atrás do complexo nuclear e sobre o rio Grataú. A fêmea foi considerada jovem por apresentar as falanges dos dígitos dos membros anteriores com ossificação incompleta. O macho foi considerado adulto. Usualmente este morcego voa em áreas abertas, clareiras na mata ou utiliza corpos d’água como rota de vôo sendo incomum encontrá-lo em áreas de vegetação densa (Wilson & LaVal, 1974; Nowak, 1993). A estimativa do tamanho das populações destas espécie de morcego através do uso de redes de neblina é incerta uma vez que morcegos insetívoros tendem a voar acima da copa das árvores em matas densas (Handley, 1967).
Família Molossidae
Molossus sp. (morcego insetívoro)
Indivíduos de espécies do gênero foram registrados através de sua observação em atividade no crepúsculo durante as coletas na mata (Tabela 71) atrás do complexo nuclear (“Trilha Telemar”). Todos os indivíduos observados estavam voando muito acima das redes e, portanto, não foram capturados.
Tabela 71 - Freqüências relativas das espécies de morcegos coletadas
| Táxon |
♂ |
♀ |
Total de indivíduos |
% |
| Artibeus lituratus |
4 |
2 |
6 |
17,14 |
| Artibeus fimbriatus |
2 |
0 |
2 |
5,71 |
| Platyrrhinus lineatus |
0 |
2 |
2 |
5,71 |
| Anoura caudifera |
4 |
0 |
4 |
11,42 |
| Lonchophylla mordax |
2 |
0 |
2 |
5,71 |
| Sturnira lilium |
1 |
2 |
3 |
8,57 |
| Carollia perspicillata |
5 |
9 |
14 |
40 |
| Myotis nigricans |
1 |
1 |
2 |
5,71 |
| Total |
19 |
16 |
35 |
100 |
Ordem Rodentia
Família Muridae
Subfamília Sigmodontinae
Juliomys pictipes (ratinho-do-mato-arborícola)
Somente um indivíduo coletado morto em ratoeira na trillha Z3. Os registros de museu indicam que esta espécie apresenta distribuição ampla na Floresta Atlântica, de onde é endêmica, embora os registros constituam-se de em geral poucos indivíduos provenientes de localidades amplamente esparsadas, em geral de áreas ainda relativamente preservadas, com pouca influência antrópica.
Oligoryzomys nigripes (ratinho-do-mato)
Dezesseis espécimes foram coletados, sendo que dois foram cariotipados. Esta espécie constitui-se em um dos roedores mais abundantes em localidades de Floresta Atlântica, podendo apresentar um padrão demográfico de explosão populacional em anos de grande produtividade primária. Em função desta característica, esta espécie tem sido registrada como transmissora de viroses silvestres, em situações em que as densidades de populações naturalmente infectadas aumentaram como resposta a um agente ambiental, e entraram em contato com populações humanas. As densidades inferidas a partir da proporção de indivíduos coletados e número de armadilhas empregadas nas linhas de coleta são congruentes com o observado em outras coletas realizadas no final da estação de chuvas em outras regiões de Floresta Atlântica.
Oryzomys nitidus (rato-do-mato)
O gênero Oryzomys é bastante complexo e diversificado. Duas espécies foram coletadas, um espécime foi coletado O. nitidus.
Oryzomys sp. (rato-do-mato)
Dois espécimes foram coletados, e o cariótipo está sendo montado para comparação com os já descritos na literatura.
Oxymycterus dasytrichus (rato-porco)
Dois espécimes foram coletados, sendo que um deles foi cariotipado. Esta espécie apresenta hábitos crepusculares e noturnos, mas freqüentemente é capturada em armadilhas dispostas durante o dia. Apresenta hábito alimentar caracteristicamente voltado para a captura de invertebrados do solo e insetos, e é capturado freqüentemente nas bordas de mata e em áreas plantadas com cana e capim próximas à mata. Por ser característico de bordas de mata, o registro de um dos espécimens em uma linha de coleta disposta em um capinzal próximo à Telemar não é um fato inusitado. Entretanto, a coleta de um dos espécimens em uma linha disposta em um bananal abandonado dentro da mata, em terreno em grande declive junto ao mar, atesta o que poderia ser o habitat mais comum desta espécie que é característica da mata atlântica, sendo que outras espécies do mesmo gênero presentes em outros biomas estão mais freqüentemente associadas à áreas de borda de matas e às regiões alagadas representadas pelas veredas e deltas.
Delomys sublineatus (rato-silvestre)
Somente um espécime foi coletado morto na ratoeira. Esta espécie tem sido mencionada como um habitante noturno e predominantemente de solo de ambientes úmidos da Floresta Atlântica. O registro desta espécie em uma localidade costeira, praticamente ao nível do mar, nesta latitude, é incongruente com as informações disponíveis na bibliografia, que só têm mencionado Delomys ocorrendo ao nível do mar nas populações mais austrais da distribuição do gênero, ou seja, entre 24 e 30 graus de latitude sul.
Akodon cursor (ratinho-do-mato).
Oito espécimens foram coletados. Espécie de hábitos noturnos, apesar de que um indivíduo foi coletado depois que a armadilha tivesse sido verificada pela manhã. Da mesma forma que Oxymycterus dasythrichus, esta espécie é também característica de bordas de mata, e encontra-se amplamente distribuída em localidades do litoral sul do Rio de Janeiro e do litoral norte de São Paulo.
Família Erethizontindae
Coendou preensilis (ouriço caixeiro)
Somente um indivíduo foi coletado, depois de ter sido encontrado atropelado na rodovia BR-101, atrás da Usina. O estado de decomposição do indivíduo não possibilitou a sexagem, e o crânio estava quebrado. Embora aparentemente comum, esta espécie torna-se rara rapidamente em ambientes sujeitos à ação antrópica.
Família Sciuridae
Sciurus ingrami (caxinguelê)
Dois indivíduos foram observados pela manhã, em dias consecutivos na mata atrás da Usina, próximo à trilha que foi usada para colocação de armadilhas. Esta espécie também está amplamente distribuída na Floresta Atlântica.
Considerações sobre os registros
Quando comparado ao número de espécies previamente levantadas na literatura para a região, onde nove ordens e aproximadamente 46 espécies de pequenos mamíferos silvestres foram registrados em um raio de 15 km da Usina Angra 3, observa-se que o número coligido na primeira campanha foi bastante inferior (20 espécies).
É interessante notar que somente uma espécie de marsupial (Didelphis aurita) foi amostrada na região no período da campanha. Embora este marsupial tenha sido uma das espécies mais abundantes na área, não foram registradas do presente estudo diversas outras especies de marsupiais que potencialmente deveriam estar presentes na área, tais como: Marmosops incanus, Chironectes minimus, Gracilinanus microtarsus, Micoureus demerarae e Monodelphis americana.
Com relação aos roedores, o número de espécies coligido na campanha (nove espécies) também se situou abaixo do esperado, pelo que pôde ser previamente levantado na literatura (25 espécies). Presume-se que um dos fatores que contribuiu para esta escassez possa ter sido a época do ano em que a campanha foi realizada, que corresponde ao final da estação de chuvas na região. Para pequenos mamiferos, marsupiais e roedores em especial, o período do ano em que as populações apresentam maiores densidades é coincidente com o final da estação seca, que na região de Angra dos Reis situa-se em setembro.
Apesar desta limitação, cabe destacar a presença nas matas situadas em torno da área de implantação do projeto de espécies aparentemente exigentes em termos da qualidade do habitat em termos da estratificação arbórea pelo menos. Este é o caso dos roedores Juliomys pictipes e Coendou prehensilis. Certamente um maior esforço de coleta proporcionaria acréscimos à presente lista, e considerando as espécies coligidas até o momento não seria surpreendente revelarem-se espécies raras ou mesmo consideradas desaparecidas na região, uma vez que a área sob influência direta do projeto tem sido poupada das modificações impostas pela presença humana, tanto no estabelecimento de lavouras como na ocupação urbana não planejada, fato que pode ser facilmente constatado pela comparação com áreas vizinhas.
7.4. Fauna Aquática - topo
O litoral do estado do Rio de Janeiro está compreendido entre as latitudes 21o S e 23o S e as longitudes 40o W e 44o W, correspondendo à porção norte da Plataforma Continental do Sudeste (PCS) do Brasil. A PCS, mais estreita em Cabo Frio - RJ (50 km) e Cabo de Santa Marta Grande - SC (70 km), apresenta a forma de lua crescente, e sua maior largura ocorre na costa de Santos - SP (230 km).
Esta área é marcada pela presença da Corrente do Brasil, que transporta águas quentes para o sul, ao longo da plataforma externa e do talude continental. Do encontro da Corrente das Malvinas, de águas frias e ricas em nutrientes, que se desloca no sentido sul-norte, com a Corrente do Brasil, aproximadamente na altura da desembocadura do rio da Prata, resulta uma massa de água denominada Convergência Subtropical que, durante o ano, apresenta deslocamentos na direção norte-sul-norte, a qual acarreta variações sazonais nas condições ambientais marinhas no sul do Brasil. Em conseqüência de tais variações, ocorrem oscilações espaciais e temporais na distribuição e abundância dos recursos marinhos vivos da região (Vazzoler, 1975).
Com relação à composição de espécies, na província Argentina, que se estende aproximadamente de Cabo Frio à Península de Valdés (Argentina), ocorre um conjunto de espécies de distribuição restrita a esta área, como também um conjunto de formas tropicais e temperadas que aí possuem seus limites geográficos, sendo considerada por Figueiredo (1981) uma região de transição faunística.
Por sua situação biogeográfica, acrescida da variedade de ambientes e recursos ecológicos passíveis de serem utilizados pela fauna de peixes, o estado do Rio de Janeiro tende a apresentar uma rica ictiofauna. Anjos (1993) destacou o potencial de biodiversidade deste trecho da costa brasileira que apresenta quatro setores com alta produtividade, os quais, a exceção de Cabo Frio, são todos relacionados a sistemas estuarinos, nas três maiores baías do estado: baías de Guanabara, Sepetiba e Ilha Grande.
A baía da Ilha Grande caracteriza-se por apresentar uma planície costeira pouco desenvolvida, uma linha de costa de traçado irregular onde se alternam pontas rochosas e pequenas enseadas, baías e ilhas. Observa-se ainda a presença de manguezais desenvolvendo-se em fundo de enseadas, em locais mais abrigados da influência de ondas.
7.4.1. Mamíferos e Quelônio - topo
7.4.1.1. Cetáceos - topo
Os cetáceos registrados no setor oeste da Baía da Ilha Grande incluem duas espécies de misticetos e quatro espécies de odontocetos. As informações reúnem os registros realizados na região de Itaorna e casos de avistagens ocasionais de baleia-jubarte, orca e golfinho-comum nas proximidades da Ilha da Gipóia, além do encalhe de um filhote de baleia-franca-do-sul em Bracuí.
Cabe ressaltar, que o número de espécies registradas neste setor da Baía de Ilha Grande pode estar subestimado, em virtude da ausência de esforços direcionados na área. Os cetáceos realizam longos deslocamentos e espécies registradas em outras áreas da Baía ocasionalmente podem utilizar esta região.
Das espécies registradas na região de Itaorna, duas possuem hábitos exclusivamente costeiros, não realizam longos movimentos e, provavelmente, utilizam a região ao longo de todo o ano: o boto-cinza (Sotalia fluviatilis) e a franciscana (Pontoporia blainvillei).
O boto-cinza, nas últimas duas décadas, tem sido alvo de inúmeros estudos, mas grande parte destas pesquisas não tem acessado com precisão parâmetros populacionais da espécie. Porém, nas regiões sul e sudeste, estudos de foto-identificação têm verificado que os indivíduos possuem elevado grau de fidelidade às respectivas áreas estudadas e este parece ser um padrão comum a outras populações. Na região de Itaorna, observações ocasionais têm mostrado movimentos diários da espécie.
A utilização da Baía de Ilha Grande e da região de Itaorna pela franciscana, espécie cujo status de conservação é definido como vulnerável, foi confirmada somente em 2001 (Azevedo et al., 2002). Tal fato tem relação com as características da espécie: pequeno tamanho do corpo, não se aproxima de embarcações em movimento e provável baixa abundância na região. Devido a estas peculiaridades da espécie, o pequeno número de registros, nesta região, pode estar relacionado à falta de estudos sistemáticos que busquem um esforço direcionado de observação e o monitoramento intensificado de encalhes e de possíveis envolvimentos da franciscana nas operações de pesca.
Azevedo et al. (2002) sugerem que a área prioritária para tal esforço está situada entre o norte de Ubatuba, no Estado de São Paulo, e o setor oeste da Baía de Ilha Grande, entre a cidade de Parati e a baía da Ribeira, Angra dos Reis, no Estado do Rio de Janeiro.
Dentre os misticetos que freqüentam a Baía de Ilha Grande a baleia-franca-do-sul, durante sua migração (meses de inverno e primavera), merece especial atenção. Este cetáceo teve seus estoques extremamente reduzidos durante a caça e a recuperação populacional, atualmente, está ameaçada pela interação com atividades humanas em águas costeiras rasas. Na Baía de Ilha Grande, pares de mãe e filhote se expõem a uma série de ameaças, que são representadas principalmente pelo emalhe em redes de pesca e o molestamento intencional. Registros de mãe e filhote desta espécie foram realizados recentemente nas praias de Mambucaba e Itaorna. O período de permanência dos indivíduos, nesta região, variou de poucos dias até uma semana. Devido às características externas (ausência de nadadeira dorsal) e ao seu comportamento de permanecer à deriva por longos períodos, o risco de choques com embarcações e enredamento em artefatos de pesca não deve ser desprezado, visto registros anteriores na região.
Na Tabela 72 estão listadas as espécies registradas na área e o respectivo status de conservação, segundo o Plano de Ação para os Mamíferos Aquáticos do Brasil (Ibama, 2001).
Tabela 72 - Lista das espécies de cetáceos ocorrentes na Baía da Ilha Grande, com seus nomes vulgares e científicos, status de conservação (Segundo Ibama, 2001) e tipo de registro.
| Nome Vulgar |
Nome Científico |
status de conservação (Ibama, 2002) |
Avistagem |
Encalhe |
| Baleia-franca-do-sul |
Eubalaena australis |
Vulnerável |
X |
X |
| Baleia-de-Bryde |
Balaenoptera edeni |
Dados Insuficientes |
X |
X |
| Baleia-minke |
Balaenoptera acutorostrata |
Dados Insuficientes |
X |
X |
| Baleia-jubarte |
Megaptera novaeangliae |
Vulnerável |
X |
X |
| Cachalote |
Physeter catodon |
Vulnerável |
|
X |
| Orca |
Orcinus orca |
Dados Insuficientes |
X |
X |
| Falsa-orca |
Pseudorca crassidens |
Dados Insuficientes |
|
X |
| Baleia-piloto-de-peitorais-curtas |
Globicephala macrorhynchus |
Dados Insuficientes |
X |
|
| Golfinho-nariz-de-garrafa |
Tursiops truncatus |
Dados Insuficientes |
X |
X |
| Golfinho-de-dentes-rugosos |
Steno bredanensis |
Dados Insuficientes |
X |
X |
| Golfinho-pintado-do-Atlântico |
Stenella frontalis |
Dados Insuficientes |
X |
X |
| Golfinho-comum |
Delphinus sp. |
Dados Insuficientes |
X |
X |
| Boto-cinza (forma marinha) |
Sotalia fluviatilis |
Dados Insuficientes |
X |
X |
| Franciscana ou toninha |
Pontoporia blainvillei |
Vulnerável |
|
X |
Dados Insuficientes - Dados conhecidos não permitem uma avaliação.
Vulnerável - Alto risco de extinção na natureza, em médio prazo.
A seguir, são apresentadas as características das 14 espécies de cetáceos registradas na Baía de Ilha Grande:
- Baleia-franca-do-sul (Eubalaena australis)
A baleia-franca-do-sul (Figura 76), espécie endêmica do hemisfério Sul, é encontrada, preferencialmente, entre as latitudes de 20º e 55º S (Jefferson et al., 1993).
Figura 76 - Baleia-franca-do-sul

Os indivíduos realizam grandes migrações latitudinais, ocupando durante o inverno e a primavera, período de reprodução, águas costeiras da costa brasileira, sendo observada do Rio Grande do Sul até o Sul da Bahia (Lodi et al., 1996).
O número de registros da espécie na costa brasileira aumentou nas últimas duas décadas, sugerindo que os estoques estão em recuperação (Santos et al., 2001). Na Baía de Ilha Grande, a espécie tem sido observada nos meses de inverno e primavera e os registros reportados na literatura confirmam que a área tem sido utilizada durante a migração. Observações de pares mãe-filhote são mais comuns, embora indivíduos solitários também sejam vistos na região (Fragoso et al., 1995). Reporta-se ainda, o encalhe de um filhote na região do Bracuí, causado por captura acidental em rede de pesca (Capistrano et al., 1996).
- Baleia-jubarte (Megaptera novaeangliae)
A baleia-jubarte (Figura 77) é encontrada em todos os oceanos e realiza longas migrações latitudinais entre as áreas de reprodução (baixas latitude) e alimentação (altas latitudes) (Jefferson et al., 1993).
Figura 77 - Baleia-jubarte

A espécie utiliza águas brasileiras para reprodução, principalmente durante os meses de inverno e primavera. A região dos Abrolhos, Sul da Bahia, é a principal área de concentração da espécie no Brasil (Siciliano, 1997).
Pares de mãe e filhote têm sido observados na Baía de Ilha Grande. Os registros reportados na literatura evidenciam que a baleia-jubarte utiliza a região durante sua migração. Assim como a baleia-franca-do-sul, o hábito costeiro expõe a espécie a diversas ameaças, especialmente o envolvimento em operações de pesca. Capturas acidentais de indivíduos adultos e filhotes têm sido reportadas para a região, sempre em redes de emalhar (Fragoso, 1995; Pizzorno et al., 1998).
- Baleia-de-Bryde (Balaenoptera edeni)
A baleia-de-Bryde (Figura 78) é o único dos balenopterídeos que não realiza longas migrações latitudinais.
Figura 78 - Baleia-de-Bryde
A espécie é encontrada em águas costeiras e oceânicas em regiões tropicais e subtropicais, não ultrapassando os 40º de latitude em ambos os hemisférios (Jefferson et al., 1993).
Muito pouco se conhece sobre a utilização do habitat e abundância da espécie na costa brasileira. Contudo, a baleia-de-Bryde parece estar presente ao longo de todo o ano costa sudeste do Brasil. Na Baía de Ilha Grande, algumas observações têm sido realizadas, incluindo relatos de predação sobre cardumes de sardinha e um encalhe na década de 80 (Fragoso et al., 1995).
- Baleia-minke (Balaenoptera acutorostrata)
A baleia-minke (Figura 79) possui distribuição cosmopolita e ocorre preferencialmente em águas oceânicas (Jefferson et al., 1993).
Figura 79 - Baleia-Minke

Na costa brasileira a espécie é raramente avistada em águas costeiras. Na Baía de Ilha Grande, os registros da espécie são raros (Hetzel & Lodi, 1996).
- Cachalote (Physeter macrocephalus)
O cachalote (Figura 80) tem distribuição cosmopolita, preferencialmente em águas oceânicas (Jefferson et al., 1993). No Brasil, a espécie é raramente avistada em águas costeiras.
Figura 80 - Cachalote

Na Baía de Ilha Grande, o cachalote foi observado em uma única oportunidade (Hetzel & Lodi, 1996).
A orca ( Figura 81) é encontrada em todos os oceanos e mares, de regiões polares até regiões equatoriais (Jefferson et al., 1993).
Figura 81 - Orca

Na costa do estado do Rio de Janeiro, a espécie parece estar presente, preferencialmente, nos meses de primavera e verão (Siciliano et al., 1999). Sua ocorrência na Baía de Ilha Grande também parece seguir esta sazonalidade, além de estar relacionada à ocupação oportunista da área em atividade de forrageamento.
Dentre os registros de Orcinus orca na Baía de Ilha Grande, destaca-se a observação de indivíduos próximos a Ilha da Gipóia e o encalhe de uma fêmea na Ilha Comprida. Os grupos observados na Baía de Ilha Grande têm sido compostos de adultos, imaturos e em alguns casos de filhotes. As poucas informações disponíveis não permitem definir o padrão de utilização da espécie na Baía de Ilha Grande, embora haja indícios de que a espécie se alimenta na região.
- Falsa-orca (Pseudorca crassidens)
A falsa-orca (Figura 82) ocorre em águas oceânicas tropicais e temperadas quentes (Jefferson et al., 1993). Devido a esta distribuição, a espécie é rara em águas costeiras. Na Baía de Ilha Grande, o único registro da espécie trata de um crânio encontrado em uma praia da região (Fragoso, 1995).
Figura 82 - Falsa-orca

- Baleia-piloto-de-peitorais-curtas (Globicephala macrorhynchus)
As baleias-piloto-de-peitorais-curtas (Figura 83) são encontradas em águas tropicais e temperadas quentes de todos os oceanos, geralmente, em áreas oceânicas (Jefferson et al., 1993).
Figura 83 - Baleia-piloto-de-peitorais-curtas
Assim como a falsa-orca, o hábito oceânico da espécie é responsável pela baixa freqüência de observação da espécie em águas costeiras. Na Baía de Ilha Grande, existe um único registro da espécie, reportado por Hetzel et al. (1994).
- Golfinho-comum (Delphinus sp.)
O golfinho-comum (Figura 84) ocorre em águas temperadas, tropicais e subtropicais de todos os oceanos (Jefferson et al., 1993). Na costa do Estado do Rio de Janeiro, a ocorrência da espécie, que tem hábitos preferencialmente oceânicos, parece estar relacionada a áreas de alta produtividade.
Na Baía de Ilha Grande, já foram reportados encalhes e avistagens do golfinho-comum (Fragoso, 1995) e a espécie parece ocorrer de forma oportunista em função da oferta de alimento e variação de fatores oceanográficos
Figura 84 - Golfinho-comum

- Golfinho-nariz-de-garrafa (Tursiops truncatus)
O golfinho-nariz-de-garrafa (Figura 85) ocorre em águas temperadas e tropicais de todos os oceanos (Jefferson et al., 1993).
Figura 85 - Golfinho-nariz-de-garrafa

A espécie possui hábitos oceânicos e costeiros, habitando saídas de estuários na região sul do Brasil (Pinedo et al., 1992).
Na Baía de Ilha Grande, encalhes e avistagens da espécie têm sido registrados (Fragoso 1995; Lailson-Brito et al., 1998), e sua ocorrência parece ser oportunista, em função da disponibilidade de alimento e fatores oceanográficos, que influenciam a distribuição da espécie.
- Golfinho-de-dentes-rugosos (Steno bredanensis)
O golfinho-de-dentes-rugosos (Figura 86) tem sido reportado como uma espécie de hábitos oceânicos, que ocorre em águas tropicais e sub-tropicais (Jefferson et al., 1993). No Brasil, entretanto, tem sido freqüentemente observado em águas costeiras (Lailson-Brito et al., 1996b).
Figura 86 - Golfinho-de-dentes-rugosos

A espécie é um dos delfinídeos mais freqüentemente registrado na Baía de Ilha Grande, seja por avistagem ou por animais encontrados mortos nas praias da região (Hetzel & Lodi, 1993; Fragoso, 1995). Apesar de estar sempre presente na região, a ausência de estudos sistemáticos não permite o conhecimento do uso da área pela espécie.
- Golfinho-pintado-do-Atlântico (Stenella frontalis)
O golfinho-pintado-do-Atlântico (Figura 87) é uma espécie endêmica do Oceano Atlântico, podendo ser encontrado em águas costeiras e oceânicas de regiões tropicais e sub-tropicais (Jefferson et al., 1993).
Figura 87 - Golfinho-pintado-do-Atlântico

Assim como o golfinho-de-dentes-rugosos, a espécie é freqüentemente registrada na Baía de Ilha Grande, seja por avistagens ou encalhes (Hetzel & Lodi, 1993; Fragoso, 1995). Contudo, não se conhece a área de vida e a utilização da área por estes golfinhos.
- Boto-cinza (Sotalia fluviatilis)
A forma marinha do boto-cinza (Figura 88) distribui-se em águas costeiras do Atlântico Ocidental desde Santa Catarina até a Nicarágua (Jefferson et al., 1993).
Figura 88 - Boto-cinza

Na Baía de Ilha Grande, o boto-cinza é o cetáceo com maior número de registros e grupos de centenas de indivíduos têm sido reportados na literatura (Lodi & Hetzel, 1999). Diferentemente de outros delfinídeos, a espécie utiliza a região ao longo de todo o ano.
Estudos pretéritos apontam para a existência de áreas preferenciais de ocorrência da espécie em algumas seções da Baía, como as proximidades de Parati e da Ribeira.
- Franciscana (Pontoporia blainvillei)
A franciscana (Figura 89) habita águas costeiras do Atlântico Ocidental, da Argentina até o Espírito Santo (Jefferson et al., 1993). A espécie é um dos cetáceos mais ameaçadas do litoral brasileiro (Ibama, 2001).
Figura 89 - Franciscana

Esta condição é devida, principalmente, à sua restrita distribuição mundial e ao hábito costeiro que a torna vulnerável aos impactos ligados às atividades humanas, especialmente a captura acidental em redes de pesca.
Apenas recentemente, a espécie foi registrada na Baía de Ilha Grande, por meio de três encalhes, dois deles ocorridos nas proximidades de Itaorna (Azevedo et al., 2001). Tais registros ampliaram os limites da distribuição da espécie para o estado do Rio de Janeiro, já que o litoral sul fluminense era considerado uma área de hiato na sua distribuição (Siciliano & Santos, 1994; Zerbini et al., 2000).
As poucas informações acerca da presença da espécie na Baía de Ilha Grande não permitem um diagnóstico mais preciso de sua situação na região. Contudo, a franciscana não costuma realizar longos movimentos e por isso a espécie deve estar presente ao longo de todo o ano nas águas da Baía de Ilha Grande.
7.4.1.2. Quelônios - topo
As tartarugas marinhas surgiram no Jurássico, sendo seu primeiro registro fóssil datado de cerca de 180 milhões de anos. São animais de vida longa que realizam migrações de centenas de milhas entre as áreas de alimentação e reprodução. Ocupam os oceanos tropicais e subtropicais, e excepcionalmente podem ser encontradas em águas frias, como no caso da tartaruga-de-couro (Dermochelys coriacea).
Algumas populações chegaram a ser compostas por milhões de indivíduos, porém hoje são poucas as populações que não estam ameaçadas pela ação humana (Sanches, 1999). O envolvimento em operações de pesca é considerado a maior causa de mortalidade de origem antrópica de tartarugas marinhas adultas e juvenis (Barata et al., 1998; TAMAR, 1999). A destruição do habitat e a coleta de ovos também ameaçam a persistência das populações (TAMAR, 1999).
Embora as tartarugas marinhas desovem em grande quantidade, as fêmeas não provêm cuidado parental e alcançam a maturidade sexual em idade avançada. Balazs (1981) estima que as tartarugas-verdes (Chelonia mydas) podem não estar reprodutivas até os 50 anos de idade. O número de ovos não é suficiente para compensar a mortalidade em várias classes etárias, causada pela caça, captura acidental em operações de pesca e perda de sítios reprodutivos. Deste modo, as populações têm declinado em todo o mundo e muitas áreas de reprodução foram extintas. Além disso, tartarugas marinhas são suscetíveis às mudanças climáticas globais uma vez que o sexo dos indivíduos é determinado pela temperatura na qual ocorre a incubação. Todas as sete espécies estão listadas no Apêndice I da CITES; seis são listados como “em perigo” ou “ameaçada” pela IUCN.
No Brasil ocorrem cinco das sete espécies de tartarugas marinhas, todas consideradas ameaçadas (TAMAR, 1999). Por exporem apenas a cabeça ou parte de seu dorso na superfície da água, as tartarugas marinhas são de difícil observação. Assim, diferentemente de cetáceos, por exemplo, que são observados mais facilmente em superfície, poucas informações são reunidas acerca dos hábitos de tais espécies na costa brasileira. A maior parte da informação está concentrada sobre aspectos ligados a reprodução, como áreas e períodos de desova.
No Sudeste, as áreas de desova localizam-se no Espírito Santo e norte do Rio de Janeiro, e as demais áreas da região são utilizadas para forrageio. Apesar dos dados não reprodutivos serem relativamente escassos, eles mostram que a costa da região Sudeste é uma importante área para o desenvolvimento do ciclo de vida das cinco espécies de tartarugas marinhas que ocorrem no litoral brasileiro (Sanches, 1999).
Os dados sobre quelônios na Baía da Ilha Grande são escassos. Contudo, a região pode ser uma importante área de alimentação para as tartarugas marinhas ocorrentes na costa brasileira, como tem sido observado para o extremo norte do litoral paulista. A região de Ubatuba é a área mais próxima que dispõe de estudos sobre as tartarugas marinhas, e possui características oceanográficas parecidas com as da Baía da Ilha Grande. Trata-se de uma importante área de alimentação, principalmente para a tartaruga-verde (C. mydas), que é a espécie mais abundante nesta área, assim como em toda a costa brasileira. No litoral de Ubatuba, as tartarugas-verdes utilizam os costões rochosos para pastoreio e repouso (Brondizio, 1994; Fernandes et al., 2002). A presença de outra espécie, a tartaruga-de-couro (D. coriacea) também foi confirmada por encalhes na Ilha Grande.
Nas áreas de alimentação, as atividades de pesca são a principal ameaça direta à sobrevivência das tartarugas marinhas. Em Ubatuba, nove diferentes artes de pesca capturam tartarugas marinhas (Brondizio, 1994; Fernandes et al., 2002).
A Baía da Ilha Grande reúne as características necessárias para sua utilização como área de alimentação das cinco espécies de tartarugas marinhas, principalmente a tartaruga-verde (C. mydas), para a qual existe uma área de alimentação conhecida próxima (Ubatuba) e com características ambientais semelhantes.
Atualmente, quase não há dados sobre a utilização da Baía da Ilha Grande pelas tartarugas marinhas, nem sobre a possível influência das atividades humanas sobre estes animais. Levando-se em consideração que as cinco espécies estão ameaçadas, reforça-se a necessidade de estudos básicos nesta região, a fim de verificar os padrões de uso da área e sua importância para as populações.
Devido às dificuldades da avistagem e identificação das tartarugas durante cruzeiros de observação, e à ausência de áreas de desova, as informações sobre a utilização da região baseiam-se na captura acidental dos animais. O monitoramento das comunidades costeiras pode fornecer informações sobre a ocorrência das tartarugas, por meio de entrevistas, e também pela notificação de animais que eventualmente se afoguem após enredamento acidental.
A seguir apresenta-se uma breve descrição das cinco espécies de tartarugas marinhas que ocorrem na Baía da Ilha Grande, baseado em Lutz & Musick (1996), Sanches (1999) e TAMAR (1999).
- Tartaruga-verde (Chelonia mydas)
Quando filhote é uma espécie onívora, tornando-se basicamente herbívora quando juvenil e adulta, podendo alimentar-se eventualmente de moluscos, esponjas e ovos de peixes. A espécie é considerada cosmopolita e as principais áreas de nidificação e alimentação estão nos trópicos. Normalmente são encontradas em profundidades rasas de até 20 m.
Figura 90 - Tartaruga-verde
- Tartaruga-cabeçuda (Caretta caretta)
A tartaruga-cabeçuda é uma espécie onívora, podendo se alimentar de crustáceos, moluscos, águas-vivas, hidrozoários, ovos de peixes e algas (Figura 91). Habitam normalmente profundidades rasas até cerca de 20 m. Quanto à reprodução das tartarugas marinhas no litoral do Brasil, observa-se que o maior número de ninhos é desta espécie.
Figura 91 - Tartaruga-cabeçuda
- Tartaruga-oliva (Lepidochelys olivacea)
A tartaruga-oliva (Figura 92)é a menor das tartarugas marinhas que ocorrem em águas brasileiras. Alimenta-se em águas mais profundas que as outras espécies, geralmente entre 80 e 100m. Porém, podem ser encontradas em águas mais rasas, principalmente em áreas próximas a estuários.
Figura 92 - Tartaruga-oliva
A espécie é onívora, alimentando-se de peixes, moluscos, hidrozoários, crustáceos, algas, briozoários, tunicados e ovos de peixe. Particularmente no Brasil possuem hábito solitário nas emergências à praia, sendo que as desovas se concentram no Estado de Sergipe.
- Tartaruga-de-pente (Eretmochelys imbricata)
Enquanto filhotes os indivíduos da espécie vivem em associação com bancos de algas do gênero Sargassum, alimentando-se principalmente de pequenos crustáceos. Na fase juvenil e adulta, a espécie torna-se onívora, podendo alimentar-se de algas, ovos de peixe, crustáceos, moluscos, ouriços, corais e esponjas. São encontradas normalmente em profundidades rasas até cerca de 40m.
A espécie pode ser mais observada em áreas tropicais, sendo as populações destas áreas compostas principalmente por subadultos. No Brasil, a principal área de desovas é o litoral norte do Estado da Bahia.
Figura 93 - Tartaruga-de-pente
- Tartaruga-de-couro (Dermochelys coriacea)
Esta espécie possui os hábitos mais pelágicos entre as tartarugas marinhas, porém pode vir alimentar-se em águas muito rasas, de até 4 m de profundidade, próximas à costa. Os hidrozoários compõem a principal parte da dieta desta espécie.
Figura 94 - Tartaruga-de-couro

Os registros são escassos e a Guiana Francesa parece ser a maior área de nidificação. As colônias no Atlântico são protegidas, e suas populações parecem estar aumentando. Na costa brasileira é a espécie mais ameaçada e reproduz-se no litoral norte do Estado do Espírito Santo e no Sul do país.
7.4.2. Ictiofauna - topo
Brum et alli (1981) foram os responsáveis pela elaboração de um relatório conclusivo e a partir deste trabalho, Furnas Centrais Elétricas (atualmente Eletronuclear) montou o Programa de Monitoração da Fauna e Flora Marinhas (PMFFM), que resultou numa série de relatórios técnicos (Furnas, 1989; 1991a; 1991b; 1992; 1993; 1994; 1995; 1996 Eletronuclear, 1997; 1998; 1999; 2000).
Alguns estudos científicos a respeito da ictiofauna marinha e/ou esturarina foram realizados na região de Angra dos Reis abordando diversos aspectos, como: levantamentos ictiofaunísticos (Brum et alii, 1981; Anjos, 1989; Hofling et alii, 1996); alimentação (Gasalla et alii, 1991; Magro, 1996; Kikuchi et alli, 1993; Gasalla & Soares, 1995); reprodução (Almeida, 1994); distribuição e abundância da comunidade íctica (Bernardes et alii, 1991; Anjos, 1993; Almeida, 1995); avaliação do setor pesqueiro (Annibal et alii, 1993; Jablonsky, 1995); ictiofauna acompanhante da pesca de camarões (Carvalho et alii, 1996). Estudos de distribuição, abundância, reprodução e alimentação de inúmeras espécies vêem sendo desenvolvidos por Andreata et alii (2002) na Baía da Ribeira e por Araújo et alii (2002) na Baía de Sepetiba.
Anjos (1987; 1993) destaca-se por ter realizado o levantamento mais completo da ictiofauna local, conforme revisão recente (Bizerril & Costa, 2001). Em seu primeiro trabalho (1987) a ictiofauna das ilhas Grande, Jorge Grego, Pombeba, Jaguanun e Itacuruça foi levantada através de observações subaquáticas e de entrevistas com pescadores locais. Já o seu segundo trabalho (1993) foi desenvolvido em uma área de 1.450 km2, que foi subdividida em 11 seções (Figura 95) correspondentes ao conjunto dos principais pesqueiros freqüentados pelos pescadores locais. Cada uma das unidades de amostragem apresenta características ambientais distintas.
Figura 95 - Principais pesqueiros da baía da Ilha Grande, segundo Anjos (1993).
Com base na similaridade taxonômica observada, Anjos (1993) identificou quatro complexos ictiogeográficos, representando os principais biótopos da baía da Ilha Grande. Desta forma, agrupam-se as unidades de Sabacú - Porcos - Abraão (ecossistemas do fundo da baía); Drago - Grego (ambientes abertos, tipicamente oceânicos); Coronel (área de substrato rochoso) e Acaia (boca da baía, com fundo arenoso e características intermediárias entre os setores internos e externos). Além de afirmar a existência de 25 grupos de peixes explotados economicamente na baía, dentre os quais destacam-se arraias, goete, linguados, pescadas e corvina que na ocasião do seu estudo perfizeram 86,9% do total das capturas realizadas.
Dentre as espécies amostradas por este autor, os linguados (grupo dominado na região pela espécie Paralichthys brasiliensis) apresentaram uma tendência de movimento sazonal dentro da baía, mostrando-se mais abundante no outono-inverno nas partes externas. As maiores capturas ocorreram na primavera-verão, nas enseadas do interior da baía.
Ainda na área de influência indireta das instalações da Central Nuclear Almirante Álvaro Alberto (CNAAA), Carvalho et alli (1996) estudaram a ictiofauna acompanhante da pesca do camarão na baía da Ilha Grande, realizando coletas nas proximidades da ponta de Acaia, Ilha da Longa e Ilha dos Porcos, onde observaram 48 espécies, sendo as da família Bothidae (linguados) as de maior freqüência de ocorrência. As espécies mais abundantes foram das famílias Gerreidae (carapicus e carapebas) e Scianidae (corvinas, papa-terra e pescadas). Estes autores registraram que a estação de coleta da Ilha da Longa é a de maior diversidade ictiofaunística.
Em estudos realizados no Saco Mamanguá (Parati), diversos autores avaliaram aspectos alimentares de distintas espécies de peixes, destacando-se:
-
Gasalla et alli (1991) analisaram os hábitos alimentares de Synodus foetens (peixe-lagarto), concluindo ser esta uma espécie piscívora predando principalmente Engraulidae (manjubas), Gerreidae (carapicus e carapebas) e Gobidae (emborés).
-
Magro (1996) estudou Diplectrumradiale (michole-da-areia) na região e concluiu serem os principais ítens alimentares desta espécie os crustáceos das famílias Caridae, Peneidae, Mysidacea e os Teleostei (peixes ósseos), variando em suas significâncias de acordo com o tamanho do predador.
-
Kikuchi et alli (1993) desenvolveram trabalhos com diferentes espécies da ictiofauna local e concluíram serem no caso de Citharichthys spilopterus (linguado) os Gobidae, Mysidacea e Caridae suas presas principais, sendo os Gobidae mais abundantes nos conteúdos estomacais dos maiores predadores, já para Achirus declivis (tapa) as principais presas seriam os Teleostei e Caridae e para A. lineatus (tapa) o principal item alimentar seria Polychaeta (vermes anelídeos bentônicos).
-
Gasalla & Soares (1995), estudaram a estrutura trófica da ictiofauna da área e concluíram existirem “indicações de que o Saco Mamanguá seja uma importante área de alimentação para peixes demersais jovens”, atribuindo-se isto ao fato de que os ambientes costeiros do referido Saco, encontram-se pouco alterados.
-
Bernardes et alii (1991) constataram que na região do Saco Mamanguá os Bothidae (linguados) e Gerreidae (carapicus e carapebas) eram as famílias mais representativas em coletas realizadas com arrastos de fundo, e Gerreidae (carapicus e carapebas) e Atherinidae (peixes-rei) em coletas realizadas com rede “picaré”.
Com relação à área de influência direta da CNAAA, Moraes et alii (1991) estudaram a ictiofauna do Saco Piraquara de Dentro (área A) e do Saco Piraquara de Fora (área B), observando índices de diversidade mais altos nos meses de primavera e verão, confirmando o que era esperado dentro de uma variação das populações de peixes destas áreas, além de registrarem diferenças acentuadas na composição das comunidades ícticas das áreas A e B.
Almeida (1994) analisou o comportamento reprodutivo de Haemulon steindachneri, coletado no Saco Piraquara de Fora, fazendo um paralelo entre o Programa Pré-Operacional e o Operacional. A espécie foi escolhida como indicador biológico para a área de necton, por ser uma das espécies de maior ocorrência e utilizada pela comunidade local na alimentação. O autor conclui que a comunidade de H. steindachneri, a nível reprodutivo, tem-se mantida equilibrada na área de influência direta da Central Nuclear de Angra dos Reis (CNAAA).
Este mesmo autor (1995) estudou a comunidade íctica do Saco Piraquara de Fora, comparando novamente a fase Pré-Operacional com os anos subseqüentes em que a Usina Nuclear Angra 1 esteve funcionando. As espécies Diapterus rhombeus, Haemulon steindachneri, Eucinostomus argenteus, Prionotus punctatus e Ctenosciaena gracilichirrhus mantiveram freqüência de 100% e foram as únicas que ocorreram nos seis períodos analisados. O estudo da diversidade específica sugeriu uma normalidade da comunidade íctica, apresentando uma tendência de aumento no máximo do índice de Shannon-Weaver ao longo dos anos estudados. Provavelmente relacionado com as oportunidades de captura.
Recentemente, durante o XXIV Congresso Brasileiro de Zoologia, realizado em Itajaí (estado de Santa Catarina), Andreata e colaboradores apresentaram uma série de trabalhos a respeito da ictiofauna da baía da Ribeira, a saber:
- Silva & Andreata (2002) - analisaram a distribuição espaço / temporal de Prionotus punctatus (cabrinha) na região, através de coletas mensais em 5 pontos amostrais, sendo 3 localizados mais internamente com características estuarinas (sob influência de aporte fluvial), onde ocorre o lançamento das águas quentes provenientes do resfriamento das usinas Angra I e II, e outro ponto mais externo no canal de entrada da baía. Sendo esta última estação a que apresentou o maior número de exemplares, seguida da estação do ponto de lançamento do efluente aquecido e dos pontos mais internos. A maior abundância foi observada durante a estação seca (inverno).
- Teixeira & Andreata (2002) - neste trabalho são indicadas as espécies de linguados da família Paralichthyidae que ocorrem na baía da Ribeira (Paralichthys brasiliensis, P. patagonicus, Etropus crossotus, E. longimanus, Syascium papillosum, Cyclopsetta chittendeni, Citharichthys arenaceus e C. spilopterus) e foram analisados aspectos reprodutivos de C. spilopterus. As amostras foram obtidas nos mesmos pontos citados em Silva & Andreata (op. cit), podendo-se observar que essa espécies apresenta um comportamento reprodutivo durante todo o ano, principalmente na primavera, e conclui-se que a espécie utiliza a baía da Ribeira como um berçário.
- Baptista & Andreata (2002) - realizaram coletas mensais em 5 pontos da baía da Ribeira para conhecer o hábito alimentar e a estratégia reprodutiva de Gerres gula, uma das espécies mais abundantes na região, com grande importância na cadeia trófica. Os itens alimentares de maior freqüência de ocorrência foram Myclitidae (Bivalva), Polychaeta, Copepoda e matéria orgânica. O pico de reprodução correu nos meses de primavera e verão.
- Baptista & Andreata (2002) - neste trabalho verificou-se a distribuição espacial por classes de comprimento na baía da Ribeira da espécie Gerres aprion. Concluíram que os indivíduos menores da espécie têm preferência pelos pontos mais interiores da baía e os maiores pelas estações externas. A baía é utilizada pela espécie como área de crescimento.
- Soares & Andreata (2002) - verificaram os itens alimentares de Genidens genidens e Bagre marinus. Para G. genidens os itens mais abundantes e freqüentes foram, respectivamente, braquiúros e escamas. Para B. marinus braquiúros e escamas foram os mais freqüentes e abundantes. A preferência alimentar destas espécies é por peixes e crustáceos.
- Freret & Andreata (2002) - analisaram os organismos que compõem a dieta alimentar de Micropogonias furnieri da baía da Ribeira. A espécie apresenta hábitos alimentares de fundo, com preferência para crustáceos (Peneídeo / Carídeo), poliquetas, escamas ctenóides e copépodas.
- Freret & Andreata (2002) - neste trabalho os autores analisaram a variação espacial e sazonal de Micropogonias furnieri, Ctenosciaena gracilicirrhus e Paralonchurus brasiliensis. M. furnieri é freqüente em estuários, onde se reproduz. As demais espécies têm características de águas mais profundas em fundo arenoso ou de cascalho, sendo capturadas na área mais externa da baía. Em relação à ocorrência sazonal das espécies, os resultados mostraram que M. furnieri ocorreu durante a estação seca (inverno), provavelmente para evitar a sobreposição de nicho em relação às demais, que ocorrem na estação chuvosa (verão).
- Longo & Andreata (2002) - os autores estudaram a distribuição espacial / sazonal, a abundância relativa e os itens alimentares de Lutjanus synagris, na baía da Ribeira. A maior ocorrência foi na estação seca (inverno). As maiores abundâncias foram nas estações mais internas, pois esses peixes são de águas quentes e rasas, sendo que os exemplares jovens preferem regiões junto à costa. Os itens alimentares identificados foram peneídeos / carídeos, escamas ctenóides e moluscos.
- Meurer & Andreata (2002) - verificaram a distribuição das espécies da família Serranidae que ocorrem na baía. Foram realizados arrastos de fundo e censo visual nos costões rochosos, sendo registradas sete espécies. Nas áreas com sedimento arenoso 6 espécies foram capturadas: Diplectrum radiale, D. formosum, Mycteroperca microlepis, M. rubra, Epinephelus niveatus e Serranus atrobranchus. Nas áreas de costão rochoso foram observadas as espécies E. marginatus, M. microlepis e M. rubra.
- Oliveira & Andreata (2002) - foram estudados os hábitos alimentares e a distribuição espacial de Diapterus rhombeus. As maiores abundâncias foram nas estações mais internas e rasas da baía. A espécie alimentou-se quase que exclusivamente de moluscos bivalves e de organismos planctônicos, seguidos por poliquetas e ofiuróides. Observaram diferenças sazonais quanto aos itens alimentares.
- Oliveira & Andreata (2002) - estudaram a variação espacial de Cynoscion jamaicensis e C. leiarchus em 5 pontos da baía. C. jamaicensis foi mais abundante na estação mais externa da baía e C. leiarchus nas mais internas.
- Vidal & Andreata (2002) - analisaram a distribuição espacial/sazonal de Dasyatis guttata e D. say. A primeira espécie apresentou preferência pelas estações mais internas, provavelmente pela oferta de alimentos onde se encontram os rios que deságuam no local e área de manguezal. A espécie D. say apresentou uma preferência pela estação que recebe águas quentes provenientes do sistema de resfriamento das usinas da CNAAA.
- Furnas (1991b) comparam os dados de freqüência por espécie, a partir de amostragens realizadas por arrastos de fundo no Saco Piraquara de Fora, nos períodos pré-operacional e operacional, não observando flutuações acentuadas nas populações de Haemulon steindachneri (cocoroca-preta), Eucinostomus gula e E. argenteus (carapicus) ao passo em que os mais acentuados decréscimos teriam ocorrido nas populações de Micropogonias furnieri (corvina), Paralonchurus brasiliensis (maria-luiza) e Selene setapinnis (peixe-galo).
Nos relatórios técnicos referentes ao necton da região, dos monitoramentos efetuados por Furnas, e mais recentemente Eletronuclear, observa-se que as espécies ocorrentes foram apresentadas em listas e a estabilidade da comunidade íctica foi inferida à partir do calculo de índices de diversidade. Sendo que por este método não foram constatadas alterações substanciais, embora seja constatado um incremento nas diversidades ao longo dos anos desde o início do funcionamento da CNAAA.
Nesses monitoramentos o método de captura “arrasto de fundo” foi largamente utilizado; o método de captura “covo” foi utilizado no período pré-operacional, de setembro de 1980 a janeiro de 1981, e no início do período operacional de outubro de 1981 a fevereiro de 1983, nas áreas A e B. A coleta do “covo” foi descontinuada devido a não padronização de amostras influenciada pelas diferentes características dos costões rochosos do Saco Piraquara de Fora; o método de coleta “rede de espera” foi utilizado nas áreas A e B, porém somente na fase pré-operacional. Cabe ressaltar que a não continuidade desta metodologia, está associado a recomendação de Brum et alii (1981) no Pré-Operacional que não considerou esta metodologia de amostragem eficiente para o estudo proposto devido ao baixo índice de captura de espécies e espécimes.
Objetivando realizar-se uma análise comparativa da Ictiofauna do Saco Piraquara de Fora (área B) nos períodos pré-operacional e operacional da CNAAA, NATRONTEC (1999) utilizou-se dos dados procedentes de amostragens realizadas por Brum et alii (1981) e pelo Laboratório de Monitoração Ambiental de Furnas / Eletronuclear, contidos nos relatórios técnicos anteriormente citados, até o período de 1995.
Dessa forma, as espécies ícticas selecionadas presentes nas amostragens realizadas por arrasto de fundo, foram classificadas quanto a seus hábitos alimentares: predador, planctófago, invertívoro e omnívoro e sua posição na coluna d’água: pelágico ou demersal. No caso de demersais, foram ainda classificadas segundo o tipo preferencial de substrato: lodoso, arenoso, arenoso-lodoso, indefinido e rochoso.
A partir dos dados referentes às abundâncias por espécies, obtidos nas amostragens realizadas desde 1980 pela UFRJ e de 1981 até 1995 por Furnas, hoje Eletronuclear, pelo método de coleta de arrasto de fundo no Saco Piraquara de Fora, foram feitas inferências sobre as flutuações nas comunidades ícticas da área de influência direta da CNAAA, sugerindo a existência de alterações na estrutura da comunidade de peixes do Saco Piraquara de Fora. Observaram-se visíveis incrementos em número, e em proporção, nas populações de peixes demersais de hábito alimentar insetivoro e omnívoro, de baixa seletividade ambiental, acompanhado de decréscimos nas populações das demais categorias, desde o início do período operacional, podendo estar associada ao ciclo das espécies. Houve uma redução na diversidade acompanhada de aumento nas capturas de predadores de grande porte e omnívoros de substratos rochosos de pequeno porte.
As alterações observadas na estrutura da comunidade de peixes do Saco Piraquara de Fora, podem estar associadas as variações naturais das populações de peixes, não sendo possível determinar a influência da CNAAA nesta comunidade, pois nos períodos analisados a usina não estava funcionando plenamente.
NATRONTEC (1999a), também, realizou observações subaquáticas nos ambientes de costão rochoso adjacentes à descarga de efluentes da CNAAA, e à marina da Eletronuclear, visando uma comparação entre as populações ícticas destes dois ambientes, os resultados apontam diferenças entre as composições das comunidades de peixes de cada ambiente amostrado. A área adjacente à descarga apresenta uma ictiofauna menos diversificada, constituída na maior parte por juvenis de espécies de baixa seletividade ambiental, além de observar-se um menor número de predadores.
Quanto aos rios, obteve-se a listagem das espécies integrante do projeto Neodat, do Museu Nacional do Rio de Janeiro, e a cobertura abrangeu os rios Bracuí, Ariró, Pereque, Grataú, Frade e Mambucaba.
Em laboratório, os peixes que a serem identificados foram transferidos de caixas de isopor com gelo para recipientes com formol a 10%. A identificação foi feita no Laboratório de Recursos Pesqueiros da Universidade Federal do Rio de Janeiro, de acordo com bibliografia específica. Alguns dos exemplares obtidos foram depositados na coleção do Laboratório de Ictiologia do Departamento de Ciências Biológicas da Universidade Estadual de Feira de Santana, na Bahia.
7.4.2.1. ctiofauna Marinha e Continental - topo
Ojetivando verificar a dominância e a participação das espécies da ictiofauna marinha e continental da área de influência de Angra 3, determinadas em campanhas anteriores, foram efetuados levantamento bibliográfico e coleta de amostras, esta apenas na área de influência direta do empreendimento (Itaorna, Piraquara de Fora e Piraquara de Dentro).
Seguindo a metodologia adotada pelo laboratório de ictiologia da Eletronuclear, foram realizados quatro arrastos demersais, com duração média de 30 minutos cada, e uma inspeção visual subaquática no costão. Para os arrastos, foi utilizada uma embarcação de pesca da frota comercial camaroneira de Angra dos Reis, equipada com rede de arrasto-com-portas de 16 m de comprimento, 1,6 m de altura, 17 m de tralha superior, malha de 20 mm e portas com cerca de 60 kg de peso.
Foram feitos quatro arrastos, sendo dois em Itaorna, um no sentido leste-oeste e o outro no sentido inverso, um em Piraquara de Fora, em direção aos túneis, que teve de ser interrompido devido a pegadores existentes no fundo que dificultaram o arrasto, e um em Piraquara de Dentro, ao longo da enseada. Os dados desses arrastos são apresentados na Tabela 73.
O material obtido por arrasto foi separado por espécie, definida quando possível no próprio local, com contagem dos indivíduos e pesagem da biomassa por espécie. Alguns exemplares das espécies não definidas no campo foram conservadas com gelo para posterior identificação em laboratório.
Tabela 73 - Informações sobre os arrastos realizados
| Discriminação |
Arrasto 1 |
Arrasto 2 |
Arrasto 3 |
Arrasto 4 |
| Local |
Itaorna |
Itaorna |
Piraquara de Fora |
Piraquara de Dentro |
| Hora inicial |
10:15 |
12:01 |
14:45 |
16:11 |
| Latitude |
23o01’32,7”S |
23o00’48,7”'S |
23o01’01,1”S |
22o59’29,6”S |
| Longitude |
044o27’11,4”W |
044o28’46,4”W |
044o26’11,6”W |
044o25’39,4”W |
| Hora final |
10:55 |
12:31 |
15:25 |
16:31 |
| Latitude |
23o01’29,”S |
23o00’47,9”S |
23o00’43,9” |
22o59’01,6”S |
| Longitude |
044o 28’25,9”W |
044o28’08,3”W |
044o26’31,2”W |
044o25’32,3”W |
| Profundidade |
10 m |
10 m |
8 m |
9 m |
| Rumo |
270o |
070o |
040o (final) |
359o |
| Extensão |
1.900 m |
1.500 m |
contornando costão |
1.000 m |
A inspeção visual subaquática foi feita por meio de mergulho, cobrindo somente o costão, em trechos situados desde as proximidades do local de captação de água em Itaorna para refrigeração do sistema dos condensadores, até o Saco Piraquara de Fora, incluindo as proximidades do local de lançamento dos efluentes.
Foram registradas 57 espécies com um total de 2.994 indivíduos coletados através de rede de arrasto-com-portas nos setores de Itaorna, Piraquara de Fora e Piraquara de Dentro (Tabela 74). Há que se considerar o caráter instantâneo, isto é do dia e horário em que se baseiam estes dados.
Tabela 74 - Numero de indivíduos e participação relativa das espécies coletadas em todos os arrastos nas áreas de influência direta da CNAAA.
Clique aqui para visualizar a tabela 74
Os dados totais dos relatórios emitidos referentes aos anos de 1997 e 1998, das duas áreas relativas a Piraquara de Fora e Piraquara de Dentro, coletados pelo Laboratório de Monitoração Ambiental da Eletronuclear, registram Diapterus rhombeus como sendo a espécie com maior abundância relativa (43,9% e 73,3%) e freqüência, seguida de Eucinostomus argenteus (8,73% e 5,5%), Haemulon steindachneri (5,7% e 3,4%) e Eucinostomus gula (3,8% e 2,2%) para estas espécies que são as indicadas para o monitoramento. Cabe mencionar que Ctenosciaena gracilicirrhus (8,63% e 5,95%) aparece em terceiro e segundo lugares para o período considerado, respectivamente, mas não está como indicadora. (ELETROBRÁS TERMONUCLEAR , 1998 e 1999).
O levantamento realizado aponta igualmente D. rhombeus como a mais abundante (46,6%), seguida de E. gula (11,0%), Micropogonias furnieri (8,0%), Larimus breviceps (5,1) e Ctenosciaena gracilicirrhus (4,9%). Por outro lado, as espécies E. argenteus e H. steindachneri não foram coletadas, no total dos arrastos realizados em Itaorna, Saco Piraquara de Fora e Saco Piraquara de Dentro.
Conforme pôde ser observado na Tabela 74, a comparação dos arrastos demonstra a clara evidência qualitativa de várias espécies de ocorrência restrita a Itaorna, fator possivelmente associado a maior dinâmica marinha dessa área.
A Figura 96 quantifica a proporcionalidade entre Itaorna e as estações do controle de monitoramento (Piraquara de Fora e Piraquara de Dentro), demonstrando as relações de maior diversidade entre o número de espécies em Itaorna e o maior número de indivíduos em Piraquaras, caracterizando a importância de utilizar a área de Itaorna como controle.
Figura 96 - Relação entre riqueza de espécies e abundância de indivíduos coletados nos arrastos em Itaorna e Sacos Piraquara de Fora e de Dentro. 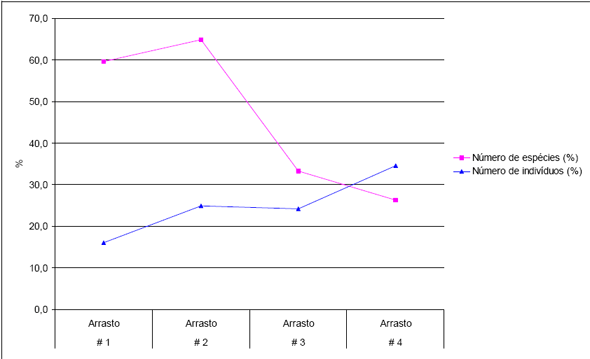
A Tabela 75 e a Tabela 76 mostram as espécies, peso por espécie, número de indivíduos e freqüência relativa dos arrastos realizados em Itaorna, sendo relevante observar, que Diapterus rhombeus e Eucinostomus gula são também comuns nessa área de referência. Outras espécies também ocorrentes, com proporções maiores que 10%, representam a diversidade das espécies da área. Cabe registrar, também, a existência de Colomesus psittacus (Família: Tetraodontidae)como primeiro registro de ocorrência para a região sudeste do Brasil.
Tabela 75 - Lista das espécies, peso por espécie, número de indivíduos e freqüência relativa do primeiro arrasto realizado em Itaorna.
Clique aqui para visualizar a tabela 75
Tabela 76 - Lista das espécies, peso por espécie, número de indivíduos e freqüência relativa do segundo arrasto realizado em Itaorna.
Clique aqui para visualizar a tabela 76
Quanto às observações subaquáticas, no costão rochoso em Itaorna, os peixes também se apresentam em maior diversidade de espécies, conforme demonstra o quadro comparativo (Tabela 77).
Tabela 77 - Lista das espécies de peixes observadas através de mergulho, em Itaorna e Piraquara de Fora e de Dentro.
| Itaorna |
Piraquara de Fora e de Dentro |
| Abudefduf saxatilis * |
Haemulon steindachneri * |
| Anisotremus virginicus * |
Eucinostomus gula * |
| Anisotremus surinamensis * |
Eucinostomus argenteus * |
| Haemulon steindachneri * |
Sphoeroides testudineus * |
| Stegastes fuscus * |
S. spengleri * |
| Mycteroperca rubra * |
Lutjanus analis |
| Diplodus argenteus * |
Scorpaena plumieri |
| Halichoeres poeyi * |
Stephanolepis hispidus |
| Prognathodes striatus |
Mugil curema |
| Pareques acuminatus |
|
| Epinephelus marginatus |
|
| Labrisomus nuchipinnis |
|
| Mugil curema |
|
| Epinephelus morio |
|
| Pomacanthus paru |
|
| Holocentrus ascensionis |
|
| Scorpaena plumieri |
|
| Stephanolepis hispidus |
|
| Scaridae |
|
Observação:espécies assinaladas com (*) foram as mais abundantes.
A região observada apresentou uma variação de profundidade entre 6 e 12 metros, com largas faixas de costão rochoso, coberto por formações coralíneas, com diversidade faunística com biocenoses incrustantes e vágeis. O fundo rochoso/coralíneo é delimitado em um fundo de areia coberto com cascalho de conchas, carapaças e fragmentos de corais.
Foi observada uma grande atividade e diversidade ictiofaunística nessa região, particularmente sobre os fundos de corais e entre as rochas, que formam grande quantidade de abrigo para os peixes. O predomínio entre os peixes, em termos de abundância, foi de Abudefduf saxatilis, Anisotremus virginicus, Anisotremus surinamensis (predominantemente jovens), Haemulon steindachneri, Stegastes fuscus, Mycteroperca rubra (predominantemente jovens), Diplodus argenteus e de Halichoeres poeyi, que ocupam amplamente essa região e são facilmente visualizados. As espécies Prognathodes striatus, Pareques acuminatus, Epinephelus marginatus, Labrisomus nuchipinnis e Mugil curema também foram observadas com boa freqüência, mas não com a abundância das primeiras. Também foram observados indivíduos de Epinephelus morio, Pomacanthus paru, Holocentrus ascensionis, Scorpaena plumieri e de Stephanolepis hispidus. Além de exemplares da família Scaridae, mas não foi possível a identificação ao nível de espécie.
De todas as espécies identificadas por observações subaquáticas apenas Diplodus argenteus foi coletado em arrasto, demonstrando que o mergulho e a identificação de visual de especialista, amplia o conhecimento sobre a ictiofauna da área, onde outros aparelhos de captura não tem capacidade de coleta.
Através do mergulho foram observadas 27% do total das espécies registradas, o que demonstra a importância complementar desse método de avaliação.
É importante frisar que os arrastos foram praticados em condições equivalentes, mas sobre setores diferentes ao que foi praticado pelo monitoramento padrão, que arrasta em trechos paralelos ao litoral em profundidades de 10/11 metros, fora da área de influência direta dos efluentes (ELETROBRÁS TERMONUCLEAR , 1999), enquanto que as coletas foram realizadas no rumo do efluente térmico, em profundidades menores (6/8 metros), portanto mais próximo do lançamento do efluente térmico no corpo d’água na enseada.
Cabe esclarecer que nessa área, em profundidades menores, devido ao substrato irregular, não é possível a prática de arrasto de fundo. A menor aproximação possível objetivou realizar coletas ictiofaunísticas em locais de uma maior influência da descarga térmica. Isto resultou na lista apresentada na Tabela 78.
Tabela 78 - Lista das espécies, peso por espécie, número de indivíduos e freqüência relativa do arrasto realizado em Piraquara de Fora.
No |
Espécies |
Peso total (kg) |
Número de indivíduos |
Freqüência relativa (%) |
| 1 |
Antennarius scaber ** |
0,20 |
2 |
0,3 |
| 2 |
Chilomycterus sp. |
0,30 |
5 |
0,7 |
| 3 |
Citharichthys arenaceus ** |
0,18 |
6 |
0,8 |
| 4 |
Citharichthys spilopterus ** |
- |
1 |
0,1 |
| 5 |
Cyclichthys spinosus * |
- |
1 |
0,1 |
| 6 |
Synodus foetens |
0,50 |
7 |
1,0 |
| 7 |
Cynoscion microlepidotus |
1,10 |
18 |
2,5 |
| 8 |
Diapterus rhombeus ** |
6,40 |
250 |
34,5 |
| 9 |
Diplectrum radiale ** |
0,80 |
19 |
2,6 |
| 10 |
Etropus crossotus ** |
0,10 |
22 |
3,0 |
| 11 |
Eucinostomus gula ** |
2,00 |
227 |
31,3 |
| 12 |
Gymnotorax ocelatus ** |
0,23 |
2 |
0,3 |
| 13 |
Gymnura sp.** |
6,8/7,4/25,0* |
3 |
0,4 |
| 14 |
Micropogonias furnieri ** |
1,85 |
143 |
19,7 |
| 15 |
Orthopristis ruber |
- |
1 |
0,1 |
| 16 |
Pomadasys curvinaeformis ** |
0,04 |
6 |
0,8 |
| 17 |
Prionotus punctatus ** |
0,35 |
12 |
1,7 |
| 18 |
Symphurus aff. Plagusia ** |
0,03 |
1 |
0,1 |
| 19 |
Trechiurus lepturus ** |
0,20 |
1 |
0,1 |
| |
Total ictiofauna |
14,08 |
725 |
100,0 |
Levando-se em conta apenas as espécies indicadoras, verificamos que D. rhombeus (34,5%) e E. gula (31,3%) dominam com uma diferença muito pequena entre elas. Em seguida, vem a espécie M. furnieri (19,7%), assim como observado no total dos arrastos e em relatórios anteriores.
Do ponto de vista comparativo, foram observadas 14 espécies comuns (**) e 57 só ocorreram no monitoramento da Eletronuclear, registradas no conjunto dos 18 arrastos anuais, realizados em freqüência bimestral, conforme descrito no relatório Eletronuclear (1999). Por outro lado, as espécies Antennarius scaber, Cyclichthys spinosus e Cynoscion microlepidotus foram coletadas nesses arrastos sem registro no relatório citado. A espécie Cyclichthys spinosus não foi observada e citada (*) em relatórios anteriores.
Quanto ao arrasto 4, realizado em área mais interna da Baía da Ribeira e mais afastada do efluente térmico, isto é, Piraquara de Dentro (Tabela 79), a amostragem caracterizou uma biomassa íctica equivalente a de Piraquara de Fora, respectivamente 15,98 e 14,08 kg, entretanto, houve uma menor diversidade (de 19 para 15 espécies) e um maior número de indivíduos (de 725 para 1037), evidenciando a presença de populações jovens, que ocorrem geralmente em áreas mais interiores da Baía da Ribeira. A presença de indivíduos jovens de Diapterus rhombeus, com freqüência relativa de 91%, evidencia a importância da espécie como indicadora para o monitoramento.
Tabela 79 - Lista das espécies, peso por espécie, número de indivíduos e freqüência relativa do arrasto realizado em Piraquara de Dentro.
| No |
Espécies |
Peso total (kg) |
Número de indivíduos |
Freqüência
relativa (%) |
| 1 |
Chaetodipterus faber * |
- |
1 |
0,1 |
| 2 |
Citharichthys arenaceus |
0,30 |
7 |
0,7 |
| 3 |
Ctaenosciaena gracilicirrhus |
0,20 |
6 |
0,6 |
| 4 |
Synodus foetens |
0,08 |
1 |
0,1 |
| 5 |
Cynoscion microlepidotus |
0,60 |
11 |
1,1 |
| 6 |
Diapterus rhombeus |
12,00 |
944 |
91,0 |
| 7 |
Diplectrum radiale |
0,15 |
1 |
0,1 |
| 8 |
Epinephelus nigritus |
0,45 |
1 |
0,1 |
| 9 |
Eucinostomus gula |
0,15 |
12 |
1,2 |
| 10 |
Genidens genidens |
0,20 |
1 |
0,1 |
| 11 |
Gymnura sp. |
0,60 |
1 |
0,1 |
| 12 |
Micropogonias furnieri |
1,00 |
42 |
4,1 |
| 13 |
Prionotus punctatus |
0,25 |
7 |
0,7 |
| 14 |
Sphoeroides greeleyi |
- |
1 |
0,1 |
| 15 |
Symphurus aff. plagusia |
- |
1 |
0,1 |
| |
Total ictiofauna |
15,98 |
1.037 |
100,0 |
A abundância relativa de jovens de Micropogonias furnieri (4,1%), espécie importante para pesca comercial e com ocorrência nos quatro arrastos demonstra a vitalidade da Baía da Ribeira. As espécies: Chaetodipterus faber, Epinephelus nigritus, Sphoeroides greeleyi só ocorreram nesse arrasto, entretanto, as duas primeiras foram registradas no conjunto de arrastos do monitoramento da Eletronuclear, enquanto Sphoeroides greeleyi e Cynoscion microlepidotus não foram registradas.
Quanto às avaliações subaquáticas, realizadas principalmente em Piraquara de Fora, observou-se um costão rochoso e estreito delimitado por fundo de areia coberto com cascalho de conchas, carapaças e pequenas rochas de granito. É uma área de baixa diversidade faunística comparada com a biocenose do costão de Itaorna, principalmente próximo aos túneis de lançamento do efluente, onde esse lançamento cria uma forte e contínua corrente.
A análise, através de mergulhos realizados em profundidades de 1,5m até 4 metros, portanto englobando a faixa de 2 metros de profundidade, onde ocorre influência da pluma térmica que contribui para uma variação média anual de 3,5 ºC (Almeida, 1995), demonstrou que esse ambiente apresenta uma reduzida diversidade ictiofaunística com participação das seguintes espécies: Haemulon steindachneri, Eucinostomus gula, Eucinostomus argenteus, Sphoeroides testudineus, que são mais abundantes, e Sphoeroides spengleri.
Lutjanus analis (juvenis), Scorpaena plumieri, Stephanolepis hispidus e Mugil curema (Tabela 75).
Em síntese as observações subaquáticas confirmam os resultados de estudos anteriores (NATRONTEC, 1999) quanto a baixa diversidade em Piraquara de Fora na proximidade do túnel de lançamento, onde se destaca, principalmente, a ausência de incrustações. Na avaliação subaquática do EIA de Angra 2 (NATRONTEC, 1999), apenas 4 espécies foram amostradas e identificadas, enquanto que na presente avaliação foram identificadas 9 espécies, sendo que Scorpaena plumieri é uma ocorrência nova para a Baía de Ilha Grande.
O levantamento da ictiofauna continental regional, conforme analisado anteriormente, compreende informações complementares, mas extremamente indiretas em relação ao eventual impacto localizado no ambiente marinho do Saco Piraquara de Fora, pois não há qualquer rio de volume significativo na área de despejo do efluente da CNAAA.
De modo a atualizar e complementar a revisão bibliográfica, obtivemos dados de registros oficiais sobre as espécies ocorrentes na região feita através de informações disponibilizadas pelo projeto NEODAT - Museu Nacional do Rio de Janeiro/UFRJ (http://www.museunacional.ufrj.br, em 27/08/2002). Os dados das principais coleções ictiológicas do mundo foram verificados quanto as localidades pertinentes ao levantamento. Foram verificados os registros de coletas para os rios Bracuí, Ariró, Pereque, Grataú, Frade e Mambucaba.
Possivelmente, algumas espécies existentes na região podem não ter sido amostradas devido a pequena quantidade de coletas direcionadas e não apareceram no levantamento. Tal fato se deve a grande quantidade de nichos oferecidos por ambientes de riachos costeiros de mata atlântica, que aumenta a quantidade de peixes especialistas, tornando mais difícil sua captura.
Como parte da fauna pertencente ao conjunto de bacias hidrográficas do leste do Brasil, muitas das espécies encontradas possuem problemas taxonômicos. Tal fato se deve à complexa e ainda pouco conhecida hidrobiologia destas bacias. A lista das espécies registradas em coleções oficiais é apresentada na Tabela 80.
Tabela 80 - Lista das espécies da ictiofauna continental registradas em coleções oficiais.
Espécies da ictiofauna continental |
| Astyanax sp. |
Poecilia vivipara (Schneider, 1801) |
| Deuterodon sp. |
Microphis brachyurus lineatus (Kaup, 1856) |
| Oligosarchus hepsetus (Cuvier, 1829) |
Geophagus brasiliensis (Quoy & Gaimard) |
| Mimagoniates microlepis (Steindachner) |
Awaous tajasica (Lichtenstein, 1822) |
| Bryconamericus sp. |
Bathygobius soporator (Valenciennes, 1837) |
| Characidium japuhybensis Travassos, 1949 |
Gobionellus shufeldti (Jordan & Eigenmann) |
| Trichomycterus sp. |
Eleotris pisonis (Gmelin, 1789) |
| Ancistrus sp. |
Centropomus parallelus Poey, 1860 |
| Rineloricaria sp. |
Mugil curema Valenciennes, 1836 |
| Schizolecis guntheri (Ribeiro, 1918) |
Rhamdia sp. |
| Kronichthys heylandi (Boulenger, 1900) |
Gymnotus pantherinus (Steindachner, 1908) |
| Pimelodella sp. |
Phalloceros caudimaculatus (Hensel, 1868) |
Considerações Finais sobre Ictiofauna Marinha e Continental
O conjunto das áreas pesquisadas, através das metodologias integradas de arrasto e mergulho, demonstraram maior diversidade de amostragens e observações, possibilitando o melhor monitoramento da ictiofauna da região. Este fato é demonstrado pelo registro de 80 espécies, que representam cerca de 45 % das espécies marinhas registradas como ocorrência na Baía de Ilha Grande.
A adição de Itaorna como área para observação da ictiofauna, possibilitou estudos comparativos e de controle sobre a área captação do efluente térmico da CNAAA.
A amostragem sobre a ictiofauna de Itaorna, que é um ambiente marinho mais aberto do que as outras áreas, representa de forma mais significativa a ictiofauna da região.
As espécies utilizadas como indicadores tradicionais foram representativas nas amostragens, entretanto considerando a maior amplitude do número de espécies registradas, poderá ser relevante pesquisar novos indicadores, principalmente associados a zona diretamente influenciada pelo lançamento do efluente termo-químico.
No monitoramento padrão, conforme mencionado em todas as conclusões dos relatórios anuais da Eletronuclear, as amostragens são realizadas fora da zona de influência, justificando também o estudo de um novo padrão para o monitoramento da ictiofauna.
Cabe ressaltar que as espécies associadas aos costões, observadas através de mergulho, que significaram 27% do total das espécies registradas, devem ser incluídas em análise comparativas futuras, pois, por estarem associadas ao litoral raso estão mais suscetíveis aos eventuais impactos causados pelo efluente.
Considerando que não há foz de rio de volume significativo na área de influência direta do lançamento, a possibilidade de impacto ambiental sobre a ictiofauna continental é praticamente inexistente.
Todos os resultados avaliados sobre a abundância e diversidade da ictiofauna demonstram que o impacto do lançamento do efluente termo-químico é extremamente localizado, necessitando de ampliarmos as avaliações futuras na área de Piraquara de Fora, ficando o monitoramento de controle em Itaorna e Piraquara de Dentro.
7.4.2.2. Ictiofauna Dulcícola - topo
A baía da Ilha Grande é um corpo de água salgada semi-confinada, com cerca de 800 km² de superfície. A semelhança das bacias contribuintes das baías de Guanabara e de Sepetiba, a bacia hidrográfica da baía da Ilha Grande, também, pode ser classificada como uma região hidrográfica. Essa bacia abrange cerca de 1.740 km² em território fluminense, compreendendo as superfícies continentais e insulares dos municípios de Angra dos Reis e Paraty, e ainda uma pequena parcela do estado de São Paulo, correspondendo ao alto curso dos rios Mambucaba, Bracuí e Ariró, este afluente do rio Jurumirim, onde estão os municípios de Bananal, Arapeí, São José dos Barreiros e Cunha (Bizerril & Primo, 2001).
A região contribuinte à baía da Ilha Grande, situada a sudoeste do estado do Rio de Janeiro, apresenta relevo acidentado e linha de litoral bastante recortada. A área da região é relativamente pequena quando comparada com a extensa linha de costa. Tal fato é explicado pela proximidade da Serra do Mar com o litoral (Cunha, 1998).
Uma característica peculiar na bacia hidrográfica da Ilha Grande é a grande quantidade de rios e córregos, que apresentam diferenças significativas entre as declividades do curso superior e o inferior, quedas de água e cachoeiras. O regime dos rios é torrencial, isto é, com grande variação nos escoamentos superficiais entre os períodos secos e chuvosos, em função da morfologia da região e dos elevados índices pluviométricos, da ordem de 2.000 mm médios anuais (os maiores do estado) - (Cunha, 1998; Bizzerril & Primo, 2001).
Quase toda a rede hidrográfica converge diretamente para o oceano, com pequenas extensões. A exceção é a bacia do rio Mambucaba, a maior da região, com nascentes no estado de São Paulo. Tem como principais afluentes, pela margem direita, os rios Guaripu e Funil e, pela esquerda, os rios Memória e Santo Antônio. Os demais rios da região com relativa importância são: Bracuí, Ariró, Mateus Nunes, Perequê-Açú, Jacuecanga, Japuíba, Pontal, Jurumirim, Bonito, Grataú, Conceição, Japetinga, Funil, Barra Grande, Pequeno, Mourisco, Meros e Paraty-Mirim (Bizerril & Primo, 2001).
O pequeno porte dos rios e o fato do baixo e médio curso destes sistemas terem sido totalmente inundados durante a transgressão marítima, gera um panorama bastante peculiar dentro do contexto da distribuição de biodiversidade no estado do Rio de Janeiro, Neste sentido verifica-se na bacia em questão uma baixa riqueza de espécies, quando comparada com as demais bacias do estado e um predomínio de formas usualmente associadas ao alto curso de rios ocupando praticamente toda a extensão dos rios, com exceção da área estuarina (Bizerril & Primo, 2001).
Na Tabela 81 é apresentada uma listagem de espécies da ictiofauna dulcícola local, com base nos poucos estudos desenvolvidos na área (Maciel, 1984; São-Thiago et alii., 1988; São-Thiago, 1990; Vianna & Caramaschi, 1990; Vianna & Caramaschi, 1991; Caramaschi et alii., 1991; São-Thiago & Caramaschi, 1992; Coutinho, 1997; NATRONTEC, 1999; Bizerril & Primo, 2001).
Na área de influência direta, Vianna & Caramaschi (1990), realizaram coletas entre setembro de 1988 e abril de 1989, pelo método de rede de espera, efetuando amostragens à intervalos de quatro em quatro horas, durante períodos de 24 horas, no trecho inferior do rio Mambucaba, objetivando analisar a partilha de recursos pelos integrantes da comunidade ictica.
Em um outro estudo, esses mesmos autores (1991), analisaram a alimentação de Deuterodon sp. no rio Mambucaba (área de influência direta), utilizando-se de redes de espera. Concluíram ser a alimentação deste peixe baseada em vegetais superiores e insetos, nesta ordem de abundância, observando a presença de areia nos conteúdos estomacais, o que sugere que o peixe busque alimento em substratos arenosos. Os autores ainda sugerem a existência de picos alimentares ao amanhecer e ao crepúsculo.
O trabalho mais recente foi realizado pela NATRONTEC (1999), com o objetivo de complementar-se as informações sobre a ictiofauna dulcícola de cinco rios da área de influência direta da CNAAA: Mambucaba, Perequê, Grataú, Frade e Bracuí. Utilizaram-se metodologias especificas as características físicas dos trechos de rio amostrados.
Observaram que o processo de adição de espécies nos rios amostrados é mais evidente nos trechos superior e médio, embora o número reduzido de amostragens realizadas nos trechos de jusante, impossibilitem conclusões definitivas. Não se registrou a presença de espécies exóticas.
O único estudo de maior duração desenvolvido na região foi o de São-Thiago (1990), que trabalhou no rio Paraty-Mirim (área de influência indireta), entre abril de 1988 e abril de 1989, analisando diversos aspectos relativos a ecologia da ictiofauna.
Para o estudo a autora utilizou-se dos métodos de coleta rede picaré, rede de espera, peneira, tarrafa, puçás e observações subaquáticas, levantando 22 espécies de peixes (Tabela 82), em 4 unidades de amostragem demarcadas ao longo do gradiente lótico, de forma que a unidade 1 corresponde ao alto curso do rio e a unidade 4, a área próxima a desembocadura.
Na distribuição das espécies, observa-se que houve aumento no número de espécies ao longo do rio, em um processo característico de adição de taxa (Shelford, 1911), ocorrendo substituição das comunidades eminentemente fluviais por arranjos com dominância de espécies marinhas.
A vazão foi identificada como um fator importante na distribuição das espécies, considerando que o seu aumento ao longo do rio acarretou em processo de adição de espécies. Esta variável parece ter tido influência na distribuição de diversas espécies, como por exemplo Characidium japuhybensis, cuja maior abundância se deu em locais com vazão entre 1,21 m³.s-1 e 2,26 m³.s-1.
Tabela 81 - Espécies de peixes nativas já assinaladas nos rios que fluem para a baía de Angra dos Reis (Bacia Hidrográfica da ilha Grande).
Clique aqui para visualizar a tabela 81
Tabela 82 - Ictiofauna amostrada no rio Parati-Mirim (Parati) e sua distribuição pelas unidades amostrais da nascente até a foz.
Clique aqui para visualizar a tabela 82
A segunda variável relevante foi a profundidade, visto que algumas das espécies ocorreram exclusivamente, ou com maior abundância, nos trechos mais rasos, enquanto outras foram mais numerosas em trechos profundos. Trichogenes longipinnis, C. japuhybensis e Trichomycterus zonatus apresentaram preferência por ambientes mais rasos. Rhamdia quelen e Gymnotus pantherinus foram mais numerosas no trecho profundo.
A presença de pedras submersas foi um aspecto identificado como relevante para alguns taxa, notadamente as espécies de Characidium, T. zonatus e T. longipinnis.
Em uma análise geral, a autora sugere 6 tipos de agrupamentos de ambientes, os quais exibem espécies que podem ser apontadas como características, a saber:
- ambientes rasos, alternando pequenas cachoeiras e patamares com pouca correnteza; pequeno volume de água; fundo de pedras; sem barranco ou com pequeno barranco: T. longipinnis, C. japuhybensis e T. zonatus;
- ambientes pouco profundos, com correnteza média; volume de água médio; fundo de pedras; pequenos barrancos marginais: Bryconamericus microcephalus e Characidium sp.;
- ambientes mais profundos, com maior correnteza; maior volume de água; fundo arenosos com pedras grandes esparsas; barranco moderado: Astyanax taeniatus, Geophagus brasiliensis, R. quelen e G. pantherinus;
- ambientes caracterizados pela presença e abundância de vegetação marginal, independentemente da correnteza, volume de água ou tipo de fundo: Schizolecis guntheri.
- ambientes com presença de poções profundos, com fundo areno-lamoso e acúmulo de folhiço: Tilapia rendalli e Oreochromis niloticus;
- ambientes de remansos, com temperaturas mais altas e pequena profundidade: Phalloceros caudimaculatus e Poecilia vivipara.
Caramaschi et alli (1991) estudaram a composição e a distribuição ictiofaunistica no rio Barra Grande em Parati, (área de influência indireta), realizando amostragens por coletas e observações subaquáticas, sugerindo serem os processos de adição e substituição de espécies, mais intensos nos trechos de jusante.
Coutinho (1997) avaliou comparativamente o impacto direto e indireto de atividades de controle de simulídeos (Diptera) utilizando formulações comerciais de larvicidas a base de Bacillus thuringiensis israelensis. As observações foram realizadas em dois rios (Pedra Branca e Muricana) na encosta da Serra do Mar, em Parati, área de influência indireta. Em relação à ictiofauna foram coletados 6.112 espécimes das espécies Bryconamericus microcephalus (Characidae), Kronichthys heylandi (Loricariidae) e Imparfinis piperatus (= I. minutus) (Pimelodidae).
Em relação aos fatores físico-químicos dos criadouros de simulídeos não se observou diferenças estatisticamente significativas entre os pontos das áreas tratadas e controle em ambos os rios. Tal observação permite a inferência que estes fatores não exercerão influência na redução da abundância da biota local.
A baía da ilha Grande possui mais de 90 ilhas, destacando-se a Grande, Gipóia e Algodão. A ilha Grande, área de influência indireta, constitui um capítulo a parte, já que reúne em seus 193 km² e 155 km de perímetro, 106 praias, além de montanhas, cachoeiras, diversos rios pequenos e duas lagoas, a do leste e do sul. Dentre os cursos de água destacam-se os córregos Itapecirica, do Sul, Parnaioca, Araçatiba, Enseada da Estrela, Abraão e Andorinha. Para as grande parte das ilhas, salvo a ilha Grande, não existem dados publicados ou mesmo acervo coletado depositado em coleções oficiais.
Maciel et alii (1984), registraram a presença de Poecilia vivipara (barrigudinho) Astyanax sp. (lambari), Mugil sp. (tainhas e paratis) e Symbranchus marmoratus (mussum) nas lagoas do Sul e do Leste, localizadas na Reserva Biológica Estadual da Praia do Sul.
Bizerril & Primo (2001) apresentam informações a respeito da ictiofauna de uma pequena área, que corresponde às bacias que fluem para a região da praia de Abraão. Nestes sistemas verifica-se uma fauna aquática que, no que se refere a seus elementos dulciaqüicolas, muito se assemelha a verificada na porção continental da macroregião da bacia da Ilha Grande. Este fato retrata a antiga conexão estabelecida, durante os períodos de regressão marítima, entre os sistemas atualmente insulares e os complexos continentais.
A lista de espécies da ictiofauna atualmente disponível é apresentada na Tabela 83.
Verifica-se um número relativamente expressivo de espécies marinhas, que ocupam grande parte dos nichos disponíveis derivado da ausência de um conjunto dulciaqüicola mais complexo (Bizerril & Primo, 2001).
Espécies como A. lineatus, G. boleosoma, G. oceanicus e D. maculatus, aparentemente restringem sua distribuição à porção final, isto é, desembocadura dos sistemas fluviais. E. pisonis e O. lineatus foram registrados até o trecho médio de rios. As demais espécies ocorrem nas porções média e superior. G. brasiliensis e P. caudimaculatus foram verificadas em praticamente toda a extensão dos rios.
Tabela 83 - Espécies de peixes registradas para a ilha Grande, Angra dos Reis, estado do Rio de Janeiro.
Táxon |
Nome Vulgar |
CHARACIFORMES
Crenuchidae
1. Characidium japuhybensis
Characidae
2. Bryconamericus microcephalus
SILURIFORMES
Pimelodidae
3. Acentronichthys leptos
4. Rhamdioglanis frenatus
5. Rhamdia quelen
Trichomycteridae
6. Trichomycterus zonatus
Loricariidae
7. Schizolecis guntheri
GYMNOTIFORMES
Gymnotidae
8. Gymnotus pantherinus
Poecilidae
Poecilinae
9. Poecilia vivipara
Cnesterodontidae
10. Phalloceros caudimaculatus
11. Phallopthychus januarius
GASTEROSTEIFORMES
Syngnathidae
12. Oostethus lineatus
PERCIFORMES
Cichlidae
13. Geophagus brasiliensis
Eleotrididae
14. Dormitator maculatus
15. Eleotris pisonis
Gobiidae
16. Awaous tajasica
17. Gobionellus boleosoma
18. G. oceanicus
PLEURONECTIFORMES
Achiridae
19. Achirus lineatus |
Canivete
Mineiro branco
Jundiá
Cambeva, moréia
Cascudinho
Sarapó
Barrigudinho
Barrigudinho
Barrigudinho
Cachimbo
Acará
Moréia, emborê
Moréia, emborê
Peixe-flor
Linguado |
7.4.3. Zooplâncton - topo
Na região da baía de Ilha Grande, o primeiro estudo sobre plâncton começou em meados da década de 40, com o levantamento realizado por Oliveira (1946) que estudou o microplâncton das Baías de Sepetiba e Ilha Grande. Neste estudo foi observada a diferença entre as regiões a leste, a oeste e a noroeste da Ilha Grande, com predominância de náuplios, copépodes e ovos de peixes, respectivamente.
Em junho de 1977, foi realizada a Operação Rio de Janeiro I, na área compreendida entre Ponta de Juatinga e Ponta Negra. Uma estação foi posicionada no interior da baía de Ilha Grande. O material coletado subsidiou uma avaliação preliminar da biomassa e da composição dos principais grupos planctônicos da região (Hubold & Gurgel, 1978), fornecendo, ainda, dados sobre a distribuição dos taliáceos (Bonecker, 1984), dos sifonóforos (Abreu & Nogueira, 1989), dos copépodes (Dias, 1994) e sobre a diversidade do zooplâncton (Bonecker et al., 1990).
Em 1980, foi realizada a Operação Rio de Janeiro II, nos meses de maio e junho, entre Ponta de Juatinga e Cabo Frio. Foram realizados três fundeios de 24 horas, com um total de 17 estações de coleta. Os copépodes foram os mais abundantes, chegando a representar 82% dos organismos da amostra. Em ordem decrescente de abundância foram: cladóceros, quetognatos, larvas de decápodes e apendiculárias. Os outros grupos representaram menos de 1% da amostra. Foi observada uma diferença espacial, com grupos associados a Água de Plataforma, como apendiculárias e hidromedusas, mais abundantes do lado oeste e grupos associados a Água Costeira, como cladóceros, mais abundantes do lado leste (Fernandes et al., 1994).
Nogueira et al. (1999) realizaram estudos sobre as comunidades do zooplâncton e ictioplâncton do Estado do Rio de Janeiro, integrando as duas operações oceanográficas (Rio de Janeiro I e II).
Durante o ano de 1980, a baía da Ribeira foi objeto de estudo detalhado com o levantamento taxonônico de sua fauna e flora marinhas, visando a obtenção de dados ambientais, em condições pré-operacionais, na região da Unidade 1 da Central Nuclear Almirante Álvaro Alberto (FUJB, 1981). A partir destes dados, Nogueira et al. (1987, 1991) realizaram um levantamento preliminar das comunidades zooplanctônicas existentes, correlacionando-as com diversas variáveis ambientais.
Em 1986, Furnas realizou uma análise estatística dos dados zooplanctônicos das coletas realizadas na fase pré-operacional, período de fevereiro de 1980 a janeiro de 1981 (Furnas, 1986).
Tendo em vista que as usinas termelétricas podem afetar os ecossistemas de várias maneiras, sendo que a principal forma seria devido ao aumento da temperatura da água, em estuários ou baías onde estariam situados os pontos de descarga dos seus efluentes (Barnete, 1972), a partir de 1986 foi iniciado um programa de monitoramento com coletas trimestrais do zooplâncton na baía da Ribeira.
Foram estabelecidos três pontos fixos: um ponto (Z3) localizado em Itaorna, na região de captação de água usada para refrigeração do circuito secundário de Angra 1 e outros dois pontos (Z4 e 47BR) na região de lançamento do efluente das Usinas Angra 1 e Angra 2, em Piraquara de Fora.
Esta estratégia amostral do monitoramento vem sendo mantida até a presente data com o objetivo de realizar um acompanhamento temporal da comunidade zooplanctônica e avaliar o impacto sobre a população de diversos organismos (Furnas, 1987, 1988, 1989, 1990, 1991, 1993, 1994, 1995, 1996, 1997, 1998, 1999, 2000 e 2001).
Em 1991, foram realizadas coletas em sete estações nas proximidades das instalações do Terminal Marítimo da baía de Ilha Grande (Tebig/Petrobras). As coletas foram em três dias consecutivos. A distribuição espacial do zooplâncton foi influenciada pela passagem da Água Central do Atlântico Sul e pela topografia da baía, onde a grande abundância de Thalia democratica modificou o gradiente vertical (Bonecker et al., 1995).
NATRONTEC (1999) realizou um levantamento de dados do zooplâncton para subsidiar o EIA de Angra 2.
Dias et al. (1999) comparou os dados coletados durante as condições pré-operacionais (1980/1981) com os levantados no período de 1991/1993, com o objetivo de avaliar a composição e variação da comunidade zooplanctônica após 10 anos de funcionamento da Usina.
A padronização da metodologia de coleta desde a campanha pré-operacional e a uniformização no tratamento das amostras, foi possível se chegar a algumas conclusões, mesmo considerando que algumas amostragens não foram realizadas devido a problemas operacionais no funcionamento contínuo da Unidade 1 da CNAAA (Furnas, 1990; 1994).
A análise das amostras referentes ao período 1986/2001 quando comparadas às obtidas em condições pré-operacionais, na região da Usina 1, nas estações Z3C (área de controle) e Z4B (área de impacto), apresentaram variações temporais semelhantes a outros sistemas costeiros tropicais. Os resultados demonstraram as influências do sistema pela contribuição de águas costeiras e oceânicas.
Comparando-se os valores atuais registrados para a densidade e a diversidade do zooplâncton com os do período pré-operacional, observou-se que houve um incremento dos mesmos, nas duas estações de amostragem, sendo estes valores compatíveis com os estudos realizados durante os últimos anos. A densidade tem apresentado, em termos gerais, um padrão de sazonalidade, maiores valores no período primavera-verão e menores no outono-inverno, acompanhando a variação dos parâmetros físicos e químicos. Na Figura 97 e na Figura 98, apresentam-se as variações da densidade do zooplâncton e diversidade específica, expressas através da padronização dos valores (valor-média/desvio padrão) com relação a um a valor médio, para todo o período de monitoramento (1980, 1986 a 2001), nas áreas de Itaorna (estação Z3) e Piraquara de Fora (estação Z4).
Desde a campanha pré-operacional até o fechamento dos estudos por parte da Universidade, foi registrada a presença de 96 táxons. Destes, 51 foram registrados no período pré-operacional. Apesar desta diferença, a comunidade zooplanctônica não sofreu modificações e mostrou-se semelhante à encontrada no período pré-operacional.
Os copépodes foram os organismos mais abundantes, seguidos dos cladóceros (principalmente Penilia avirostris), das larvas de moluscos, das apendiculárias e das larvas de poliquetos. As espécies de Copepoda Paracalanus quasimodo, Parvocalanus crassirostris, Temora stylifera, Oithona hebes (espécies costeiras e estuarinas), Oncaea minuta e Oncaea curta (espécies com preferência por águas mais salinas) e de Cladocera Penilia avirostris (espécie costeira), são as mais abundantes e freqüentes.
Figura 97 - Variações da densidade do zooplâncton e diversidade específica, para todo o período de monitoramento (1980, 1986 a 2001) na área de Itaorna (estação Z3).
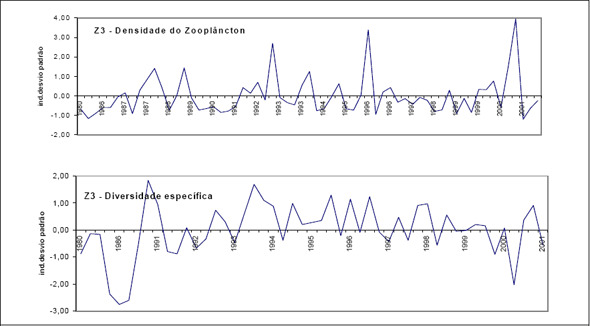
Figura 98 - Variações da densidade do zooplâncton e diversidade específica, para todo o período de monitoramento (1980, 1986 a 2001) na área de Piraquara de Fora (estação Z4).
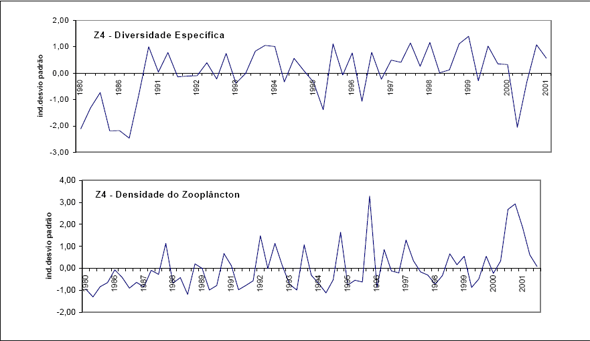
Os resultados demonstram as influências sofridas pelo sistema por parte da presença de águas costeiras e estuarinas, principalmente, e oceânicas. Como a influência de águas mais salinas às vezes só pode ser observada pela presença de espécies, tais como Oncaea venusta, Oncaea minuta, Oncaea curta, Acrocalanus longicornis, Oithona plumifera, Macrosetella gracilis (copépodes), Krohnitta pacifica e Caecosagitta macrocephala (quetognatos), reforça a importância dos parâmetros físico-químicos serem amostrados juntamente com a coleta de dados biológicos.
Os organismos zooplanctônicos encontrados na baía da Ribeira são característicos de regiões costeiras e estuários do litoral brasileiro, sendo semelhantes as associações planctônicas encontradas em alguns estudos como no estuário do rio Igarassú (Paranaguá et al., 1979); no estuário do rio Botafogo, em Recife (Paranaguá e Nascimento-Vieira, 1984); na baía de Ilha Grande (Bonecker et al., 1995), na baía de Sepetiba (Coelho-Botelho et al., 1999) e baía da Guanabara (Nogueira et al., 1989; Valentin et al., 1999; Schutze & Ramos, 1999).
As amostras de zooplâncton foram obtidas por arrasto vertical com rede cônico-cilíndrica de 200 µm de abertura de malha, 60 cm de diâmetro de boca e 2,0 m de comprimento. Para o estudo da distribuição espacial foram coletadas amostras em doze pontos, sendo seis em Piraquara de Fora e seis em Itaorna.
Para o estudo da microvariação temporal foi feita uma amostragem de alta freqüência em duas estações fixas - Z3 e Z4 -, com coleta de seis em seis horas, em maio de 2002.
Para avaliação qualitativa e quantitativa dos crustáceos decápodes foram utilizadas as amostras obtidas pela rede bongô dos arrastos oblíquos. Para melhor interpretação dos dados, as amostras da malha de 330 µm foram utilizadas para estimar a densidade e a composição desse grupo, empregando-se as amostras da malha de 500 µm como complemento do inventário faunístico.
Para a estimativa do volume de água filtrada foi acoplado, entre o aro e o centro da boca da rede, um fluxômetro (modelo General Oceanics) previamente aferido.
Imediatamente após a coleta, o material foi fixado e conservado em formaldeído diluído a 4% em água do mar e tamponado com tetraborato de sódio à razão de 20g.L-1.
Em laboratório, os organismos zooplanctônicos foram contados e triados sob microscópio estereoscópico Aus Jena. Na triagem, as amostras foram fracionadas com o auxílio do Folsom Plankton Sample Splitter (McEwen et al., 1954), sendo os organismos triados até um limite mínimo de 100 indivíduos (Frontier, 1981) por grupo. Para a triagem de Copepoda utilizou-se o método de subamostras com subamostrador de volume conhecido (10 mL).
A densidade dos organismos foi calculada em função do volume de água filtrada pela rede e expressa em ind.m-3. O volume de água filtrada foi calculado pela fórmula:
V = a.n.c
onde: V= volume de água filtrada (m3)
a= área da boca da rede (m2)
n= número de rotações (rot)
c= fator de aferição do fluxômetro (m.rot-1)
A identificação dos grupos zooplanctônicos Copepoda, Cladocera, Appendicularia, Salpidae, Doliolidae, Chaetognatha e larvas de Decapoda, importantes na caracterização da comunidade local, foi feita sob microscópio estereoscópico Zeiss Stemi SV6 e microscópio Olympus Tokyo modelo 210582. A identificação das espécies em níveis taxonômicos inferiores foi feita com o auxílio de bibliografia especializada (Copepoda: Björnberg, 1981, e Bradford-Griece, 1999; Cladocera: Ramírez, 1981, e Onbé, 1999; Larvas de Decapoda: Boltovskoy, 1981 e 1999, e Williamson 1957 e 1982; Doliolidae: Esnal, 1999a; Salpidae - Esnal, 1999b;
Appendicularia - Esnal & Daponte, 1999; Chaetognatha - Mc Lelland, 1980, e Mc Lelland, 1989). Os demais grupos foram apenas quantificados.
7.4.3.1. Caracterização do Zooplâncton - topo
Após a análise das amostras referente as radiais de Itaorna (área controle) e Piraquara de Fora (área de impacto), assim como da amostragem de microvariação em pontos fixos (Z3 e Z4), foram encontrados 27 grupos zooplanctônicos, os quais são apresentados na Tabela 84, onde os grupos e espécies que estão representadas com (*) ocorreram pela primeira vez na área de estudo.
Este número foi superior aos que foram observados durante o estudo pré-operacional e ao longo do monitoramento (1986 a 2002). Esta diferença deveu-se a ocorrência dos grupos Foraminifera, Radiolaria, Turbelaria, Cumacea e Salpidae, não assinalados anteriormente, a identificação das larvas de decápodes e ao aumento do esforço amostral.
Tabela 84 - Relação dos grupos taxonômicos do zooplâncton identificados.
Clique aqui para visualizar a tabela 84
Os grupos zooplanctônicos Ctenophorae e Heteropoda (Mollusca), coletados durante o estudo Pré-Operacional e ao longo do monitoramento (1986 a 2002), não foram registrados na presente amostragem.
Distribuição Espacial (radiais)
Na radial de Itaorna as densidades do zooplâncton variaram entre 31.568 ind.m-3 (estação R7) e 261.364 ind.m-3 (estação R9) - (Tabela 85) e nas radias de Piraquara de Fora a variação foi de 31.603 ind.m-3 (estação R1) e 291.295 ind.m-3 (estação R3) - (Tabela 86).
Tabela 85 - Densidade dos grupos e espécies zooplanctônicas (ind.m-3) das amostras da rede de 200 µm dos arrasto horizontal na área de Itaorna.
Clique aqui para visualizar a tabela 85
Tabela 86 - Densidade dos grupos e espécies zooplanctônicas (ind.m-3) da malha de 200 mm dos arrastos horizontais da área de Piraquara de Fora.
Clique aqui para visualizar a tabela 86
De uma maneira geral, a distribuição espacial apresentou os maiores valores nas estações mais afastados dos pontos de captação e descarga do efluente da usina (Figura 99).
Figura 99 - Distribuição espacial da densidade do zooplâncton (ind.m-3) nas áreas de Itaorna (a) e Piraquara de Fora (b) da campanha de maio de 2002.
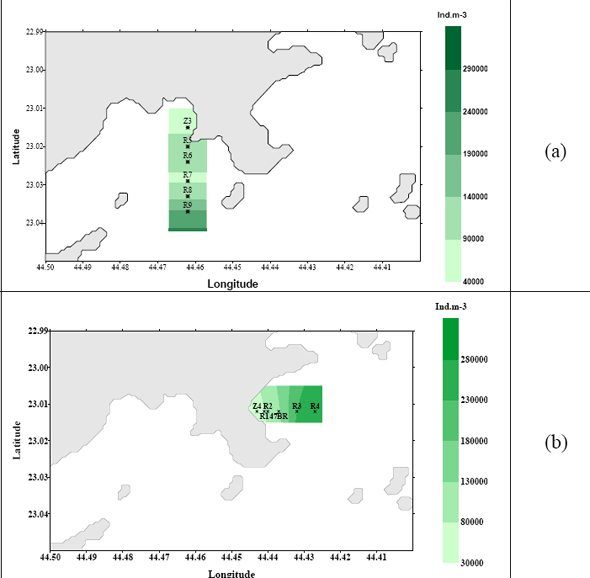
A dominância dos grupos foi semelhante para as duas áreas. Os copépodes foram os mais abundantes, com 75% do total do zooplâncton em Itaorna e 80% em Piraquara de Fora. Os cladóceros foram o segundo grupo mais abundante com percentuais de 20% e 18% em Itaorna e Piraquara de Fora, respectivamente. Os demais grupos zooplanctônicos foram representados em outros (Figura 100).
Figura 100 - Percentual (%) dos principais grupos zooplânctônicos obtidos nas áreas de Itaorna (a) e Piraquara de Fora (b).
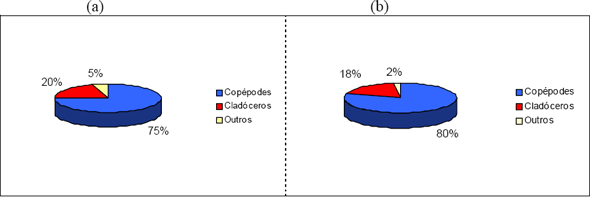
A dominância de Copepoda em número de espécies (composição) e densidade (abundância) em todas as estações de coleta é um fato comum no zooplâncton marinho.
Em Itaorna, foram registrados 27 táxons de copépodes (Tabela 85). Jovens e adultos da ordem Calanoida foram os mais abundantes, com 66% da densidade total. Das 17 espécies identificadas Temora stylifera (30%), Paracalanus quasimodo (15%), Clausocalanus furcatus (6%) e Corycaeus giesbrechti (6%) foram as mais representativas.
A diferenciação encontrada entre as estações, foi devido a ocorrência de jovens da Ordem Calanoida encontrados nas duas primeiras estações (estação Z3R e R5) e na última estação da radial de Itaorna. Pode-se observar, a partir da estação R5, a elevação da densidade da espécie Corycaeus giesbrechti assim como a ocorrência das espécies Macrosetella gracilis, Sapphrina nigromaculata, Oncaea venusta, Candacia curta (espécies com preferência por águas mais salinas), Pseudodiaptomus acutus, Labidocera fluviatilis e Subeucalanus pileatus (espécies com características costeiras e estuarinas) o que caracteriza a mistura da água costeira e de plataforma nestas estações. A ocorrência da espécie Undinula vulgaris, indicadora da presença da Corrente do Brasil, nas três últimas estações da radial (estações R7, R8 e R9) confirma a presença de águas oceânicas na região.
Em Piraquara de Fora foram registrados 26 táxons de copépodes (Tabela 86). Os jovens e adultos da ordem Calanoida também foram os mais abundantes (58%). Das 22 espécies identificadas, Temora stylifera (42%), Corycaeus giesbrechti (12%), Paracalanus quasimodo (5%) e Oithona plumifera (3%) foram as mais abundantes.
Em relação a abundância de espécies, houve uma diferenciação nas espécies dominantes (Temora stylifera, Corycaeus giesbrechti, Paracalanus quasimodo e Oithona plumifera), entre as estações da radial. Temora stylifera foi a espécie dominante em cinco das seis estações de amostragem, principalmente nas duas últimas estações (R3 e R4). A exceção deste padrão de dominância foi observada na estação 47BR, onde a espécie Corycaeus giesbrechti foi a mais abundante. Pode-se observar que a ocorrência das espécies com características costeiras e estuarinas, como por exemplo, Centropages furcatus, Pseudodiaptomus acutus e Oithona hebes ficaram restritas as quatro primeiras estações da radial (Z4R, R1, R2 e 47BR). Foi observada, também, a ocorrência da espécie Oithona plumifera (quarta em termos de abundância) e Oncaea venusta, espécies com preferência por águas mais salinas, o que caracteriza a mistura da água costeira e de plataforma nestas estações.
A fauna de Copepoda das duas radiais é formada por espécies epipelágicas sendo dominada por organismos neríticos, seguidos de espécies com preferência por águas mais salinas. As espécies encontradas foram registradas anteriormente no estudo Pré-Operacional e ao longo do monitoramento (1986 a 2002). Foi ainda observada uma maior ocorrência de espécies características de águas mais salinas, seguindo o padrão encontrado ao longo deste ano (Furnas, 2002 a e b).
Ressalta-se a diferenciação encontrada entre as duas radiais. As espécies Undinula vulgaris, Subeucalanus pileatus, Candacia curta, Sapphrina nigromaculata e Macrosetella gracilis só foram encontradas na área de Itaorna, enquanto que Paracalanus aculeatus, Calocalanus contractus, Oithona hebes, Oithona plumifera, Oncaea curta, Copilia mirabilis, Microsetella norvergica e Clytemnestra scutellata só foram encontradas na área de Piraquara de Fora.
Os branquiópodos (cladóceros) foram representados por quatro espécies: Pseudevadne tergestina, Penilia avirostris, Pleopis schmackeri e P. polyphemoides. A última espécie ocorreu somente em Piraquara de Fora.
Pseudevadne tergestina é freqüentemente encontrada em associação com Penilia avirostris em águas quentes sem ultrapassar, em ambos hemisférios, a latitude de 40ºS (Ramírez, 1981). P. avirostris é uma espécie cosmopolita, encontrada em regiões neríticas e marcadamente eurihalina (Ramírez, 1981). Valores elevados de temperatura parecem influenciar o desenvolvimento desses organismos. O caráter euritérmico e eurihalino desta espécie, associado à capacidade de reproduzir-se por partenogênese, permite-lhes explorar rapidamente os mais diversos biótipos, desde que ricos em alimento (Björnberg, 1981). A espécie P. schmackeri é geralmente encontrada em águas costeiras quentes (Onbé, 1999). Esta espécie foi encontrada pela primeira vez na área de estudo. P. polyphemoides é uma espécie cosmopolita e eurihalina, mencionada para ambientes com alto grau de diluição, como lagunas salobras, mares internos com influência continental e setores internos de portos (Ramírez, 1981).
Em Itaorna, a densidade de decápodes variou entre 1.849 ind.100m-3 (estação Z3R) e 4.950 ind.100m-3 (estação R9) - (Tabela 87) e em Piraquara de Fora a variação foi de 4.570 ind.100m-3 (estação R3) a 10.134 ind.100m-3 (estação R4) - (Tabela 88).
Tabela 87 - Densidades (ind.100m-3) dos decápodes planctônicos, larvas e pós-larvas (megalopas) encontradas nas amostras coletadas com malha de 330 µm, em arrastos oblíquos, na área de Itaorna.
Táxons/Estações |
Z3R |
R5 |
R6 |
R7 |
R8 |
R9 |
| Penaeidae |
14 |
0 |
72 |
11 |
29 |
43 |
| Lucifer faxoni |
162 |
1341 |
1504 |
844 |
1291 |
1479 |
| Sergestidae |
7 |
38 |
0 |
16 |
15 |
14 |
| Thalassinidae |
0 |
0 |
57 |
64 |
160 |
57 |
| Callianassidae |
122 |
89 |
344 |
32 |
0 |
28 |
| Upogebidae |
165 |
32 |
229 |
170 |
203 |
142 |
| Caridea |
317 |
221 |
387 |
361 |
232 |
441 |
| Alpheidae |
101 |
253 |
201 |
297 |
653 |
341 |
| Oplophoridae |
7 |
0 |
14 |
0 |
44 |
14 |
| Hippolytidae |
22 |
108 |
158 |
244 |
145 |
341 |
| Lysmata wurdemanni |
22 |
0 |
0 |
48 |
29 |
71 |
| Albuneidae |
0 |
0 |
29 |
0 |
0 |
0 |
| Paguridae |
22 |
32 |
0 |
127 |
87 |
71 |
| Porcellanidae |
7 |
25 |
14 |
21 |
44 |
57 |
| Leucosiidae |
0 |
6 |
14 |
11 |
11 |
14 |
| Majidae |
22 |
16 |
11 |
0 |
0 |
21 |
| Pinnotheridae |
7 |
13 |
25 |
42 |
11 |
21 |
| Portunidae |
0 |
66 |
36 |
21 |
22 |
43 |
| Hexapanopeus schmitti |
5 |
9 |
4 |
11 |
11 |
28 |
| Mennipe nodifrons |
0 |
0 |
4 |
0 |
4 |
0 |
| Megalopa de Brachyura |
14 |
9 |
14 |
16 |
7 |
14 |
| Não identificado |
0 |
0 |
7 |
11 |
0 |
0 |
| Total |
1849 |
3137 |
4669 |
3820 |
4657 |
4950 |
Tabela 88 - Densidades (ind.100m-3) dos decápodes planctônicos, larvas e pós-larvas (megalopas) encontradas nas amostras coletadas com malha de 330 µm, por arrastos oblíquos, na área de Piraquara de Fora.
| Táxons/Estações |
Z4R |
R1 |
R2 |
47BR |
R3 |
R4 |
| Penaeidae |
264 |
0 |
186 |
0 |
0 |
0 |
| Lucifer faxoni |
240 |
565 |
291 |
1078 |
1964 |
1973 |
| Sergestidae |
29 |
88 |
0 |
0 |
0 |
82 |
| Thalassinidae |
0 |
0 |
0 |
0 |
0 |
41 |
| Callianassidae |
294 |
0 |
0 |
0 |
34 |
41 |
| Upogebidae |
0 |
351 |
372 |
517 |
135 |
822 |
| Caridea |
1938 |
2018 |
2327 |
2888 |
271 |
3124 |
| Alpheidae |
176 |
1272 |
1396 |
733 |
220 |
1644 |
| Oplophoridae |
29 |
0 |
0 |
0 |
68 |
0 |
| Hippolytidae |
646 |
1755 |
2234 |
1336 |
1033 |
740 |
| Lysmata wurdemanni |
0 |
44 |
93 |
86 |
169 |
41 |
| Paguridae |
235 |
395 |
419 |
86 |
254 |
493 |
| Porcellanidae |
0 |
0 |
0 |
43 |
0 |
0 |
| Grapsidae |
88 |
175 |
116 |
0 |
17 |
41 |
| Majidae |
147 |
132 |
23 |
32 |
42 |
123 |
| Pinnotheridae |
2349 |
1601 |
2024 |
830 |
237 |
822 |
| Portunidae |
147 |
88 |
0 |
22 |
25 |
41 |
| Hexapanopeus schmitti |
352 |
0 |
140 |
108 |
102 |
92 |
| Mennipe nodifrons |
0 |
526 |
0 |
0 |
0 |
0 |
| Megalopa de Brachyura |
88 |
22 |
23 |
11 |
0 |
0 |
| Não identificado |
0 |
0 |
70 |
11 |
0 |
10 |
| Total |
7025 |
9032 |
9715 |
7780 |
4570 |
10134 |
Em Itaorna, foram encontrados 21 táxons de decápodes (Tabela 87). A infraordem Penaeoidea contou com 46% da abundância e foi melhor representada pelo grupo holoplanctônico, Lucifer faxoni (44%), alcançando a densidade de 1.504 ind.100m-3 na estação R6. Os carídeos (camarões de estalo) foram o segundo grupo mais abundante, com 34% da densidade total nas amostras. Dentre os Brachyura, os Portunidae (siris) foram os mais abundantes (2%), sendo representada pelas espécies Hexapanopeus schmitti e Mennipe nodifrons. Foi encontrada também a família Pinnotheridae que são pequenos caranguejos que vivem em associação com esponjas e anêmonas.
Em Piraquara de Fora, foram encontrados 21 táxons de decápodes (Tabela 88). Os carídeos (camarões de estalo) representaram 54% da densidade total nas amostras. A família Hyppolitidae, com a espécie Lysmata wurdemanni,representou 17% e a família Alpheidae representou 11%. Os braquiúros (siris e caranguejos) foram o segundo em termos de abundância com 22%. A infraordem Anomura obteve 4% da abundância, sendo representada principalmente pelos paguróideos, cuja densidade variou de 86 a 493 ind.100m-3.
Dentre os quetognatos Flaccisagitta enflata foi a espécie mais abundante, seguida por Ferosagitta hispida e Sagitta bipunctata nas amostras de Itaorna. As duasúltimas espécies foram registradas pela primeira vez para a área de Itaorna. Em Piraquara de Fora, Parasagitta friderici foi a espécie mais abundante, seguida por F. enflata e S. bipunctata. Segundo Costa (1971), F. enflata é uma espécie epiplanctônica de águas quentes, sendo reconhecidamente a segunda espécie de Chaetognatha mais abundante, nas baías dessa porção da costa brasileira. P. friderici é descrita como uma espécie nerítica e epiplanctônica, que prefere águas de baixas salinidades próximas, mas é capaz de tolerar salinidades oceânicas (Mc Lelland, 1980). Esta espécie é comum e bem representada nas baías e enseadas da costa sudeste brasileira. Krohnitta pacifica é considerada boa indicadora de águas quentes de superfície (Dadon & Boltovskoy, 1982), além de indicadora de mistura recente de água oceânica com água costeira (Pierce & Wass, 1962). S. bipunctata é encontrada em áreas pelágicas tropicais e subtropicais. A espécie Ferosagitta hispida é considerada endêmica no Atlântico, sendo abundante em áreas neríticas tropicais e subtropicais, assim como P. tenuis (Boltovskoy, 1981b).
As apendiculárias apresentaram seus valores de densidade oscilando entre 180,22 ind.m-3 (estação Z3R) e 3.288,85 ind.m-3 (estação R9) em Itaorna e entre 39,85 ind.m-3 (estação Z4R) e 1.399,32 ind.m-3 (estação R4) em Piraquara de Fora.
Foram identificadas quatro espécies de apendiculárias(Oikopleura dioica, Oikopleura rufescens, Oikopleura fusiformis e Oikopleura longicauda) em Itaorna. Para Piraquara de Fora, além das quatro já citadas ocorreu, ainda, a espécie Fritillaria sp. O. dioica foi a espécie mais abundante nas duas áreas. Esta espécie é típica de regiões neríticas e da Água Costeira e Tropical. O. rufescens é considerada abundante até o paralelo de 45ºS (Esnal, 1981 e Esnal & Daponte, 1999) e O. longicauda está associada a águas quentes com ampla distribuição no Atlântico Sudoeste (Esnal et al., 1999). O. fusiformis é considerada termófila e cosmopolita (Esnal, 1981), sendo registrada pela primeira vez na área de estudo.
Dentre os Thaliacea, a família Doliolidae foi pouco representativa nas duas áreas. Doliolum nationalis foi mais frequente que Dolioletta gegenbauri, ocorrendo em todas as estações nas duas áreas. D. gegenbauri ocorreu pela primeira vez em Itaorna e teve sua ocorrência assinalada pela segunda vez para Piraquara de Fora, tendo sido registrada anteriormente durante o outono do corrente ano (Furnas, 2002b). Estas espécies são cosmopolitas de águas quentes e comumente encontradas ao longo de todo o litoral brasileiro.
Foi encontrada pela primeira vez, na área de Itaorna, a espécie Thalia democratica (Família Salpidae), com densidade de 0,04 e 0,12 ind.m-3, nas estações R6 e R9, respectivamente. Esta espécie é considerada por Barth (1964) como indicadora de Água Tropical. É cosmopolita, característica de águas quentes, sendo a mais abundante da família Salpidae.
Microvariação Temporal
As densidades totais do zooplâncton na área de Itaorna foram superiores aos valores encontrados para Piraquara de Fora (Figura 101). Em Itaorna a variação foi de 106.176 ind.m-3, na coleta das 06:17 horas durante a maré vazante, a 193.281 ind.m-3, na coleta das 16:50 horas também na maré vazante (Tabela 89), enquanto que em Piraquara de Fora os valores de densidade variaram de 21.966 ind.m-3, na coleta das 17:55 horas durante a maré vazante, a 40.455 ind.m-3, na coleta das 12:05 horas na maré enchente (Tabela 90). Não foram observadas variações em relação às situações de maré.
Figura 101 - Variação da densidade do zooplâncton (ind..m-3) durante a amostragem de microvariação nas áreas de Itaorna (estação Z3) e Piraquara de Fora (estação Z4).
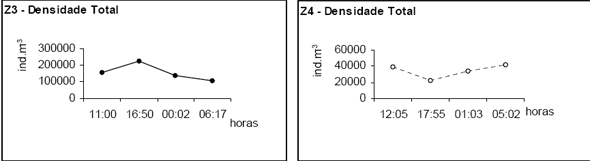
Tabela 89 - Densidade dos grupos e espécies zooplanctônicas (ind.m-3) encontradas nas amostras coletadas com a rede de 200 µm em arrasto horizontal durante a amostragem de microvariação em Itaorna. Maré enchente/alta nos períodos de 11:00-11:30 e 00:02-00:29. Maré vazante/baixa nos períodos de 16:50-17:15 e 06:17-06:46
Clique aqui para visualizar a tabela 89
Tabela 90 - Densidade dos grupos e espécies zooplanctônicas (ind.m-3) encontradas nas amostras coletadas com a rede de 200 µm em arrasto horizontal durante a amostragem de microvariação em Piraquara de Fora. Maré enchente/alta nos períodos de 12:05-12:46 e 01:03-01:40. Maré vazante/baixa nos períodos de 17:55-18:37 e 05:02-05:35.
Clique aqui para visualizar a tabela 90
Os copépodes foram os mais abundantes com densidades superiores a 90% do total do zooplâncton, nas duas áreas de estudo. Houve uma inversão na densidade de copépodes entre as duas áreas.
Em Itaorna, o menor valor ocorreu na maré vazante das 06:17 horas e o maior valor foi na maré vazante das 16:50 horas, enquanto que em Piraquara de Fora o menor valor foi na maré vazante das 17:55 horas e o maior ocorreu na maré vazante das 05:02 horas (Figura 102). Em Piraquara de Fora, apesar de ter apresentado menores densidades, foram registrados mais táxons (35) de copépodes do que em Itaorna (21). A dominância dos copépodes (Calanoida) foi a mesma das radiais.
Figura 102 - Variação dos valores de densidade de Copepoda (ind.m-3) durante a amostragem de microvariação nas áreas de Itaorna (estação Z3) e Piraquara de Fora (estação Z4).
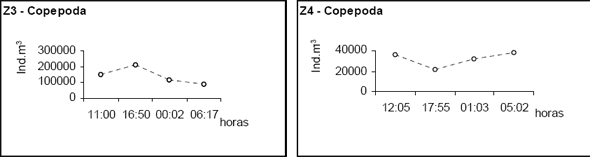
Jovens e adultos da Ordem Calanoida foram os mais abundantes, com 84% da densidade total em Itaorna e 73% em Piraquara de Fora. A comunidade de copépodes foi semelhante nas duas áreas variando a quantidade de espécies encontradas e o percentual entre elas. Das 17 espécies identificadas em Itaorna as mais representativas foram Clausocalanus furcatus (31%), Paracalanus quasimodo (9%) Temora stylifera (7%) Oncaea venusta (3%) e Corycaeus giesbrechti (2%). Em Piraquara de Fora foram identificadas 24 espécies, sendo as mais representativas as espécies Temora stylifera (20%), Paracalanus quasimodo (12%), Corycaeus giesbrechti (7%), Clausocalanus furcatus e Oithona plumifera (6%).
Em Itaorna foram observadas espécies neríticas (Acartia lilljeborgi e Calanopia americana) e com preferência por águas mais salinas (Undinula vulgaris e Corycaeus speciosus), indicadoras da Corrente do Brasil. Além destas, Copilia mirabilis, Calocalanus contractus e Oithona plumifera só foram coletadas durante a maré enchente/alta, enquanto que Euterpina acutifrons, só foi coletada durante a maré vazante/baixa. Já em Piraquara de Fora observou-se espécies neríticas (Subeucalanus pileatus e Oithona oculata), espécies com preferência por águas mais salinas (Acrocalanus longicornis e Clytemnestra scutellata) e representantes da Ordem Monstrilloida que só foram coletadas durante a maré enchente/alta, enquanto que Euterpina acutifrons, Oithona similis (espécie com preferência por águas mais frias) e Oncaea minuta só foram coletadas durante a maré vazante/baixa.
Os cladóceros (Branchiopoda) também apresentaram seus valores de densidade invertidos nas duas áreas. Em Itaorna a variação foi de 432 ind.m-3 na maré enchente das 11:00 horas, a 79.612 ind.m-3 na maré vazante das 16:50 horas. Em Piraquara de Fora a densidade variou de 703 ind.m3 na maré vazante das 17:55 horas a 2.455 ind.m-3 na maré enchente das 12:05 horas (Figura 103).
Figura 103 - Variação dos valores de densidade de Cladocera (ind.m-3) durante a amostragem de microvariação nas áreas de Itaorna (Z3) e Piraquara de Fora (Z4).
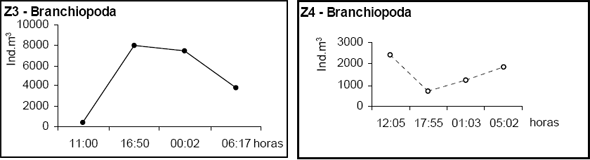
Além das quatro espécies identificadas em Piraquara de Fora (Penilia avirostris, Pseudevadne tergestina, Pleopis polyphemoides, P. schmackeri)foi encontrada mais uma (Podon intermedius) em Itaorna. P. avirostris foi a espécie mais abundante seguida por P. tergestina. P. polyphemoides, encontrada nas amostras das radiais de Piraquara de Fora,foi registrada pela primeira vez para a área de Itaorna.
As maiores densidades dos decápodes ocorreram em Piraquara de Fora. A estação 1Z3 apresentou uma grande quantidade de ctenóforos prejudicando a análise da amostra. Não foi observado um padrão de distribuição dos decápodes em relação as situações de maré (Figura 104).
Figura 104 - Variação dos valores de densidade das larvas de Decapoda (ind.100m-3) durante a amostragem de microvariação nas áreas de Itaorna (Z3) e Piraquara de Fora (Z4).
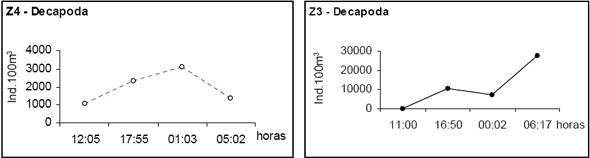
Foi encontrado um total de 17 táxons de decápodes em Itaorna (Tabela 91), nas marés enchente/alta de 11:00-11:30 e 00:02-00:29 e vazante/baixa de 16:50-17:15 e 06:17-06:46. Em Piraquara de Fora, foi encontrado um total de 18 táxons de decápodes (Tabela 92), nas marés enchente/alta de 11:30-12:05 e 01:03-02:29 e vazante/baixa de 16:50-17:55 e 05:02-06:17.
Tabela91 - Densidades (ind.100m-3) das larvas de decápodes das amostras da malha de 330 µm, em arrasto oblíquo, na amostragem de microvariação em Itaorna.
Táxons/Estações
Horário das coletas |
1Z3
11:30 |
2Z3
16:50 |
3Z3
02:29 |
4Z3
06:17 |
| Penaeidae |
0 |
0 |
14 |
0 |
| Lucifer faxoni |
534 |
118 |
415 |
528 |
| Sergestidae |
0 |
14 |
28 |
16 |
| Callianassidae |
41 |
0 |
84 |
10 |
| Upogebidae |
54 |
97 |
338 |
140 |
| Alpheidae |
159 |
374 |
225 |
186 |
| Hippolytidae |
50 |
83 |
0 |
103 |
| Lysmata wurdemanni |
18 |
42 |
0 |
16 |
| Hippidae |
2 |
0 |
0 |
5 |
| Paguridae |
27 |
194 |
352 |
47 |
| Paguridae Megalopa |
0 |
0 |
0 |
16 |
| Porcellanidae |
5 |
0 |
28 |
0 |
| Leucosiidae |
5 |
3 |
7 |
0 |
| Majidae |
25 |
59 |
53 |
62 |
| Pinnotheridae |
0 |
308 |
176 |
103 |
| Portunidae |
7 |
3 |
18 |
5 |
| Hexapanopeus schmitti |
2 |
17 |
130 |
47 |
| (Megalopa de Brachyura) |
2 |
21 |
18 |
18 |
| Totais |
1081 |
2303 |
3108 |
1373 |
Tabela 92 - Densidades (ind.100m-3) das larvas de decápodes , encontradas nas amostras da malha de 330 µm, em arrasto oblíquo, na amostragem de microvariação em Piraquara de Fora.
Táxons/Estações
Horário de coleta |
1Z4
12:05
|
2Z4
17:55
|
3Z4
01:03
|
4Z4
05:02 |
Penaeidae |
0
|
240
|
71
|
147 |
Lucifer faxoni |
0
|
1321
|
3064
|
7731 |
Sergestidae |
0
|
240
|
189
|
589 |
Callianassidae |
0
|
0
|
71
|
74 |
Upogebidae |
0
|
800
|
189
|
2356 |
Alpheidae |
0
|
320
|
283
|
3166 |
(Megalopa Alpheidae) |
0
|
0
|
24
|
0 |
Palemonidae |
0
|
0
|
24
|
0 |
Oplophoridae |
0
|
0
|
94
|
0 |
Hippolytidae |
0
|
2801
|
471
|
2651 |
Lysmata wurdemanni |
0
|
80
|
24
|
0 |
Paguridae |
0
|
0
|
613
|
1546 |
Paguridae Megalopa |
0
|
40
|
377
|
0 |
Porcellanidae |
0
|
0
|
24
|
0 |
Leucosiidae |
0
|
0
|
0
|
221 |
Majidae |
0
|
200
|
283
|
1399 |
Pinnotheridae |
0
|
480
|
719
|
4271 |
Portunidae |
0
|
40
|
12
|
295 |
Hexapanopeus schmitti |
0
|
320
|
271
|
884 |
(Megalopa de Brachyura) |
0
|
120
|
212
|
810 |
(Juvenil de Brachyura) |
0
|
0
|
24
|
0 |
Totais |
0
|
10365
|
7342
|
27612 |
Em termos gerais, a dominância dos decápodes foi a mesma nas duas áreas de estudo. Os carídeos (camarões de estalo) foram os mais abundantes, representando mais de 30% do total. O segundo em abundância foram os Luciferídeos (Lucifer faxoni) com mais de 20% da abundância total. Em Piraquara de Fora as densidades de larvas de decápodes foram superiores.
Os valores de densidade de quetognatos em Itaorna (Z3) variaram de 6,29 ind.m-3 na estação realizada às 06:17 horas e 74,24 ind.m -3 , coletados durante a maré vazante de às 16:50 horas. Em Piraquara de Fora se observou uma inversão na distribuição dos valores de quetognatos (Figura 105), com a menor densidade encontrada nas amostras realizadas às 17:55 horas, durante a maré vazante (1,77 ind.m -3 ) e a maior densidade (3,01 ind.m -3 ) encontrado na amostragem realizda às 12:05 horas.
Figura 105 - Variação dos valores de densidade de Chaetognatha (ind.m-3) durante a amostragem de microvariação nas áreas de Itaorna (Z3) e Piraquara de Fora (Z4).
A composição das espécies dos quetognatos encontrados foi semelhante às encontradas nas radiais, com exceção da presença de Mesosagitta mínima e Krohnitta subtilis que foram registradas pela primeira vez para a área de Itaorna e Caecosagitta macrocephala registrada pela primeira vez em Piraquara de Fora.
Mesosagitta minima é característica de regiões de plataforma até 45º S, enquanto Krohnitta subtilis é considerada uma espécie transoceânica de regiões tropicais e subtropicais (Furnestin, 1957; McLelland, 1980; Boltovskoy, 1981b). Caecosagitta macrocephala é uma espécie oceânica, rara em águas costeiras (Mc Lelland, 1989), já tendo sido assinaladas para a região (Almeida Prado, 1961; Nogueira et al., 1987; Dias et al., 1999).
Tanto para as apendiculárias (Figura 106) como para os doliolídeos (Figura 107) as maiores densidades foram encontradas em Itaorna. As espécies dominantes destes dois grupos foram, respectivamente, Oikopleura dioica (Appendicularia) e Doliolum nationalis (Doliolidae). Estas espécies também dominaram nas amostras das radiais.
Figura 106 - Variação dos valores de densidade de Appendicularia (ind.m-3) durante a amostragem de microvariação nas áreas de Itaorna (Z3) e Piraquara de Fora (Z4).
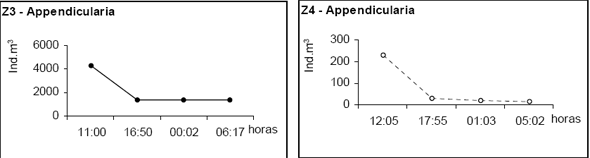
Figura 107 - Variação dos valores de densidade de Doliolidae (ind.m-3) durante a amostragem de microvariação nas áreas de Itaorna (Z3) e Piraquara de Fora (Z4), na campanha de maio de 2002.
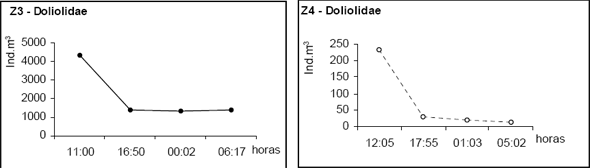
Considerações finais sobre o Zooplâncton
Após a análise das amostras referentes a distribuição espacial, assim como as amostragens de microvariação temporal, pode-se demonstrar as influências sofridas pelo sistema por parte da presença de águas costeiras e estuarinas, principalmente, e águas oceânicas. Estas influências são semelhantes a outros sistemas costeiros tropicais.
Foi identificado um total de 27 grupos zooplanctônicos, número superior ao observado durante o estudo Pré-Operacional e ao longo do monitoramento que vem sendo realizado (1986 a 2002). Os copépodes foram os organismos mais abundantes, seguidos dos cladóceros (representados, principalmente, pela espécie costeira Penilia avirostris) e das larvas de Mollusca (Gastropoda e Bivalvia).
Espacialmente, as estações mais distantes dos pontos de captação e saída do efluente foram as que apresentaram maiores densidades.
Ao longo dos ciclos de marés, as densidades totais de zooplanctôn foram superiores em Itaorna, mas com um menor número de espécies. Apesar de não ter sido observado diferença na composição do zooplâncton ao longo do ciclo de maré foi observado diferença na densidade entre Itaorna e Piraquara de Fora. As tendências de caracterizações ao longo dos ciclos de maré foram opostas, evidenciando um comportamento diferenciado em função, provavelmente, de padrão de circulação nas duas áreas.
A fauna de copépodes foi formada por espécies epipelágicas sendo dominada por organismos neríticos (Temora stylifera, Paracalanus quasimodo e Corycaeus giesbrechti), seguidos de espécies com preferência por águas mais salinas (Clausocalanus furcatus, Oithona plumifera e Oncaea venusta). A maioria das espécies encontrada no presente estudo foi registrada anteriormente no estudo pré-operacional e ao longo do monitoramento (1986 a 2002), sendo que o aumento, tanto do número de táxons quanto da abundância de espécies características de águas mais salinas, tem sido observado, principalmente, ao longo deste ano (2002).
O padrão de dominância das espécies sofreu uma ligeira alteração, devido a ocorrência de algumas espécies com preferência por águas mais salinas entre as mais abundantes. Este fato indica a mistura recente de água oceânica com água costeira, fato não observado nos parâmetros físicos e químicos, onde a salinidade registrada foi sempre inferior a 33. Outras espécies com preferência por águas mais salinas foram encontradas em menor densidade como Undinula vulgaris (espécie indicadora da Corrente do Brasil), Microsetella norvergica, Microsetella rosea, Macrosetella gracilis, Sapphirina nigromaculata, Copilia mirabilis, Clytemnestra scutellata (Copepoda), Caecosagitta macrocephala, Krohnitta pacifica (Chaetognatha), Thalia democratica (Salpidae), Doliolum nationalis e Doliolleta gegenbauri (Doliolidae). A comunidade zooplanctônica não sofreu modificações e mostrou-se semelhante às coletas anteriores e a encontrada no período pré-operacional.
7.4.4. Ictioplâncton - topo
O ictioplâncton constitui um componente de grande importância dentro da comunidade planctônica, especialmente as meroplanctônicas (Ciechomski, 1981). Isto deve-se ao fato da grande maioria dos peixes teleósteos possuem ovos planctônicos, e que estas espécies se caracterizam por uma fecundidade muito elevada. Além disso, as larvas e pós-larvas das espécies que possuem ovos demersais, presos ao substrato, levam uma vida pelágica, integrando-se a comunidade planctônica (Ciechomski, op. cit.).
Os estuários, baías, lagoas e águas costeiras desempenham um papel importante no ciclo vital de algumas espécies de peixes, funcionando como áreas de desova e recrutamento de suas larvas (Weiss & Muelbert, 1980; Schawamborn & Bonecker, 1996, entre outros). Estes se agrupam nestas áreas e se beneficiam com a possibilidade de abundância de alimento e proteção de predadores. Mais de 70% de juvenis de peixes de espécies de importância comercial no Atlântico Norte-Ocidental, têm uma parte de sua vida associada a sistemas estuarinos (Clark, 1967; Mc Hugh, 1967).
O Estado do Rio de Janeiro apresenta três importantes baías: Guanabara, Sepetiba e Ilha Grande. Estas baías estão submetidas a distúrbios ecológicos por receberem vários tipos de poluição: portuária, industrial e urbana. A baía de Ilha Grande é um sistema estuarino, parcialmente misturado, com riachos e manguezais em seu contorno e um total de 365 ilhas e ilhotas (Bonecker et al., 1995). Pelas condições ecológicas e paisagísticas, as ilhas constituem um considerável potencial turístico e de lazer. Apesar da intensa pesca na região, muito pouco se conhece sobre a composição da comunidade ictioplanctônica.
Os primeiros estudos sobre a distribuição e abundância de ovos e larvas de peixes na área de estudo foram realizados pelo Instituto Oceanográfico da Universidade de São Paulo, na década de 70. Richardson (1959) investigando a sardinha na costa brasileira verificou que os comprimentos desses peixes, desembarcados em Santos, eram menores e suas gônadas estavam em estágio de maturação, enquanto os desembarcados no Rio de Janeiro eram maiores e prontos para desovarem. Devido a essa afirmação, Matsuura (1971) iniciou a coleta quantitativa de ovos e larvas de sardinha na região de Ilha Grande, entre os anos de 1969/1970. Foram realizados cinco cruzeiros, na área de distribuição da sardinha, de Cabo Frio à ilha de São Sebastião, sendo que três deles foram realizados entre Barra de Guaratiba e ilha de São Sebastião, com seis estações de coleta na baía de Ilha Grande.
A partir dos dados coletados, Matsuura (1971) realizou um estudo sobre a distribuição e abundância dos ovos de sardinha, com o objetivo de analisar a fase inicial do seu ciclo de vida e verificar o potencial reprodutivo na costa sul do Brasil. Além da descrição dos ovos, consta a determinação dos períodos de desova anual (setembro a março) e diário (pouco antes da meia-noite).
Foram realizados dois cruzeiros entre Cabo de São Tomé e Cabo de Santa Marta Grande, nos anos de 1970/1971 e 1971/1972, dando continuidade aos estudos de distribuição e abundância de ovos de Sardinella brasiliensis (Matsuura, 1975a). Apenas o cruzeiro de 1971/1972 apresentou uma estação no interior da baía de Ilha Grande, tendo sido realizada, ainda, uma comparação desses dados com os dos cruzeiros anteriores.
Utilizando estas mesmas amostras Matsuura (1975b) fez uma descrição da morfologia das larvas de Sardinella brasiliensis a fim de determinar um padrão para a identificação das mesmas.
No mesmo ano, Matsuura (1975c) realizou um estudo das correntes superficiais da área de desova da sardinha, através de cartões-de-deriva. O autor levantou a hipótese da corrente do Brasil não influenciar diretamente o transporte e distribuição de ovos e larvas de S. brasiliensis na região.
Reunindo os três cruzeiros (1969/1970; 1970/1971 e 1971/1972), Matsuura (1977) estudou a distribuição e abundância de larvas de peixes da sardinha verdadeira (Sardinella brasiliensis). Apenas uma estação nos dois últimos cruzeiros foi posicionada no interior da baía de Ilha Grande.
Durante o período de 1975 a 1978, foram realizados sete cruzeiros oceanográficos na região sudeste, com o intuito de avaliar a potencialidade de peixes pelágicos e estudar a influência das condições oceanográficas sobre a reprodução de peixes (Matsuura et al., 1980). Destes cruzeiros, somente uma estação estava posicionada no interior da baía de Ilha Grande. Do material coletado, os ovos e larvas foram triados e utilizados para ampliar o conhecimento sobre os estágios larvares de várias espécies, resultando em teses e publicações (Matsuura, 1977; Matsuura & Katsuragawa, 1981; Matsuura & Sato, 1981; Nakatani, 1982; Matsuura & Yoneda, 1986; Sato & Matsuura, 1986; Kurtz, 1992). Contudo, esses estudos não destacam as estações e sim áreas, perdendo-se, portanto, toda a informação da estação do interior da baía.
Com a construção da Central Nuclear Almirante Álvaro Alberto (CNAAA) na baía de Ilha Grande foi realizado um levantamento da fauna e flora em três áreas da baía da Ribeira, no período de fevereiro de 1980 a janeiro de 1981, obtendo-se ao todo 144 amostras de plâncton. O ictioplâncton foi pouco representado nas amostras coletadas durante o ano, apresentando uma abundância relativa menor que 0,2% do total de organismos zooplanctônicos coletados. Foram encontradas duas ordens (Perciformes e Pleuronectiformes) e cinco famílias (Engraulidae, Clupeidae, Gobiidae, Sparidae e Serranidae). Os engraulídeos foram os mais freqüentes, ocorrendo ao longo de todo o ano, seguidos dos gobídeos. As sardinhas (Clupeidae) ocorreram de fevereiro a maio e de outubro a janeiro (FUJB, 1981a).
Após esse levantamento preliminar, somente em 1986 foram retomadas as coletas de zooplâncton, incluindo a abundância relativa dos ovos e larvas de peixes. O plano amostral foi reduzido, entretanto a metodologia de coleta, com arrasto vertical, foi mantida. Os ovos e larvas de peixes continuaram sendo subamostrados e foram classificados apenas como ovos e larvas, não sendo identificados adequadamente, pois o Programa de Monitoração da Fauna e Flora Marinha da CNAAA não contempla a análise do ictioplâncton (Furnas, 1987, 1988, 1989, 1990, 1991, 1993, 1994, 1995, 1996; Eletronuclear 1997, 1998, 1999, 2000 e 2001).
Em 1991 foram realizadas, em três dias consecutivos, coletas em sete estações próximas ao Terminal da baía de Ilha Grande (Tebig/Petrobras). As coletas foram realizadas durante a passagem de uma frente fria. Nas águas superficiais mais frias, ocorreu uma elevada densidade populacional de salpas (Thalia democratica) modificando o padrão de distribuição das espécies. A presença de T. democratica causou um impacto negativo à comunidade ocasionando a redução ou ausência dos ovos e larvas de peixes. As espécies de ovos de peixes mais frequentes foram Engraulis anchoita (16%) e Harengula jaguana (8%). Foram identificadas larvas das famílias: Engraulidae, Clupeidae e Blenniidae (Parablennius pilicornis) (Bonecker et al., 1995).
Dias et al. (1999) realizaram um estudo sobre a variação na estrutura da comunidade zooplanctônica próxima a CNAAA em dois ciclos anuais (1980/1981 e 1991/1993), com dez anos de diferença. O ictioplâncton foi melhor representado no segundo período de estudo. Apesar da alta frequência, os ovos e larvas de peixes apresentaram baixa abundância relativa. Este fato está associado ao tipo de arrasto realizado. Os arrastos verticais não são apropriados para o estudo do ictioplâncton.
No EIA, realizado pela NATRONTEC (1999), para a implantação da Usina de Angra 2, o ictioplâncton foi relacionado como classe Osteichthyes.
Geralmente o ictioplâncton não é a fração mais numerosa do meroplâncton, sendo as larvas de crustáceos e moluscos as mais abundantes. Entretanto, os ovos e larvas de peixes podem representar cerca de 10% do total de indivíduos coletados (Bonecker et al., 1990).
A abundância dos ovos e larvas de peixes encontrados, até o último relatório do monitoramento da Central Nuclear Almirante Álvaro Alberto (CNAAA), não chegam a representar 1% do total da comunidade zooplanctônica.
O ictioplâncton, apesar de estar inserido dentro da comunidade zooplanctônica, necessita de cuidados durante a coleta, de modo que sua amostragem possa ser representativa. Os programas de amostragem de ictioplâncton, quando eficazes, são um poderoso instrumento para ajudar a determinar: as áreas e as épocas de desova, o crescimento e a mortalidade das formas larvares, as relações tróficas entre as larvas de peixes e o zooplâncton, além de outros (Unesco, 1975).
Os arrastos realizados para o estudo do ictioplâncton, no monitoramento da CNAAA, foram do tipo vertical. Este tipo de arrasto ocasiona uma subamostragem do ictioplâncton, principalmente das larvas de peixes, pois elas "percebem" a rede e escapam, devido a sua mobilidade (Phonlor, 1973).
As recomendações mais aceitas para a coleta do ictioplâncton com objetivo de determinar a distribuição e a abundância destes, tanto no espaço como no tempo, é a utilização de uma rede do tipo bongô, com malhas de 330 e 500 µm e arrastos oblíquos. A rede deve ser equipada com um fluxômetro para que se possa estimar o volume de água filtrada pela rede. O volume filtrado não deve ser menor que 100 m³ (Unesco, 1975).
Segundo Haedrich (1983), a ocorrência de espécies costeiras em estuários deve-se ao fato desses ambientes apresentarem características semelhantes às águas costeiras. As famílias importantes encontradas nesses ecossistemas, em zonas tropicais são: Clupeidae, Engraulidae, Mugilidae, Sciaenidae, Gobiidae e Achiridae. Na baía de Ilha Grande foram encontradas três famílias (Engraulidae, Clupeidae e Gobiidae) das citadas por Haedrich (op. cit.). Além dessas, foram encontradas também as famílias Sparidae, Serranidae e Blenniidae. Nos estuários brasileiros são encontradas outras famílias, muito comuns na nossa costa, que ainda não foram citadas para a baía de Ilha Grande, tais como: Sciaenidae, Blenniidae, Gerreidae, Achiridae, Paralichthyidae e Tetraodontidae. Essas espécies são citadas por Weiss & Muelbert (1980) para a Lagoa dos Patos, por Sinque et al. (1982) para o complexo-estuarino de Paranaguá, por Faro & Bonecker (2000) para a baía de Guanabara e por Castro & Bonecker (1996) para o estuário do rio Mucuri.
Pela listagem de peixes adultos encontrados desde o levantamento preliminar (FUJB, 1981b) observa-se que muitas espécies são típicas de ambientes estuarinos, desovando nesses ambientes.
Muito pouco se conhece sobre o ictioplâncton da baía de Ilha Grande. São citadas, até o momento, três espécies, seis famílias e duas ordens na região. Outras regiões estuarinas da costa brasileira, tanto ao norte como ao sul da região, apresentam uma variabilidade muito maior.
Em virtude de sua importância social e paisagística, sugere-se a elaboração de um programa de estudos específicos sobre o ictioplâncton. O estudo de ovos e larvas de peixes possibilita o conhecimento das áreas de desova e recrutamento de larvas de peixes que são explorados comercialmente, particularmente em relação a pesca comercial e esportiva, prática muito comum neste importante sistema do sul fluminense.
A amostragem de ictioplâncton foi feita por distribuição espacial e por microvariação temporal, utilizando-se no primeiro caso seis pontos de coleta em duas radiais, localizadas em Itaorna e Piraquara de Fora. Para avaliar a microvariação do plâncton foram feitas coletas de 6 em 6 horas, durante 24 horas, em dois pontos fixos, sendo um em Itaorna e o outro em Piraquara de Fora.
A coleta foi feita por arrasto oblíquo, no sentido superfície-fundo-superfície, com duração média de 10 minutos, tendo sido utilizadas uma rede bongô, de dois aros de 60 cm de diâmetro de boca, e duas redes cilíndrico-cônicas, de 250 cm de comprimento e aberturas de malha de 330 e 500 mm. As redes foram equipadas com fluxômetro (General Oceanics) previamente aferidos, para estimar o volume de água filtrada. As amostras foram acondicionadas em frascos de polietileno e imediatamente fixadas com formaldeído a 4%, preparado com água do mar, e tamponadas com tetraborato de sódio à razão de 20 g.l-1.
Em laboratório, os ovos e as larvas de peixes foram triados sob microscópio estereoscópico (Aux-Jena). As amostras que apresentaram uma grande quantidade de ovos de peixes foram fracionadas com o emprego de Folsom Sample Splitter (Mc Ewen et al., 1954). O número de indivíduos coletados foi extrapolado para um volume padrão (100 m3) pela da expressão:
y = (x/V).100
onde:
y = número de ovos ou larvas por 100 m3
x = número de ovos ou larvas coletados na amostra
V = volume de água filtrada
O volume de água filtrada pela rede foi obtido pela fórmula:
V = a.n.c
onde:
V = volume de água filtrada (m3)
a = área da boca da rede (m2)
n = número de rotações (rot)
c = fator de aferição do fluxômetro (m.rot-1)
Para a identificação do ictioplâncton foi utilizado microscópio estereoscópico Zeiss modelo Stemi SV6. A identificação das larvas de peixes foi realizada com auxílio de bibliografia especializada (Leis & Rennis, 1949; Fahay, 1983; Moser, 1996 e outros). As larvas que estavam destruídas, impedindo a identificação, foram agrupadas em não identificadas. O inventário faunístico foi baseado na classificação de Nelson (1994).
7.4.4.1. Caracterização do Ictioplâncton topo
Dos ovos e larvas de peixes coletados em Itaorna e Piraquara de Fora foram identificados um total de 19 famílias e 20 espécies (Tabela 93). Os táxons assinalados com (*) já haviam sido citados para a regiâo da baía de Ilha Grande nos trabalhos de dados anteriores. A maioria dos ovos (94%) não foi identificada por não apresentar características possíveis de diagnóstico, sendo identificado somente os ovos da família Engraulidae.
Tabela 93 - Composição do ictioplâncton da área de Itaorna e Piraquara de Fora.
Clique aqui para visualizar a tabela 93
A família Serranidae (Diplectrum sp.) ocorreu somente nas amostras coletadas com o objetivo de verificar a distribuição espacial e as famílias Sparidae (Archosargus rhomboidalis), Dactyloscopidae (Dactyloscopus sp.) e Tetraodontidae ocorreram somente nas amostras de microvariação.
As famílias de peixes encontradas no presente estudo são todas costeiras e comuns em baías e estuários brasileiros. A maioria dos táxons já foi registrada na baía do Espírito Santo (ES) (Bonecker et al., 1991), sistema estuarino do rio Mucuri (BA) (Castro & Bonecker, 1996) e baía de Guanabara (RJ) (Bonecker, 1997; Castro, 1998).
A grande variedade de larvas de peixes coletadas numa área restrita da baía de Ilha Grande (Praia de Itaorna e Saco Piraquara de Fora), em uma única campanha, indica a importância da área como local de desova e recrutamento para algumas espécies e reflete a grande diversidade ambiental que caracteriza este ecossistema.
Distribuição Espacial
Durante a campanha de campo, nas amostras coletadas em rede bongô nas 12 estações das áreas de Itaorna e Piraquara de Fora, observou-se o seguinte: as densidades de ovos de peixes da malha de 330 mm variaram de 63,07 a 839,26 ovos.100 m-3 (Tabela 94), com os valores mais altos na área de Itaorna, todos acima de 100 ovos.100 -3 (Figura 108); na malha de 500 mm, os valores de densidade variaram de 21,83 a 551,00 ovos.100 m-3, apresentando uma inversão da dominância, com os maiores valores registrados em Piraquara de Fora (Figura 109).
Tabela 94 - Valores de volume, número e densidade de ovos de peixes e de larvas coletados em Itaorna e Piraquara de Fora.
Clique aqui para visualizar a tabela 94
Figura 108 - Distribuição espacial da densidade de ovos de peixes da malha de 330 mm da área de Piraquara de Fora (a) e Itaorna (b).
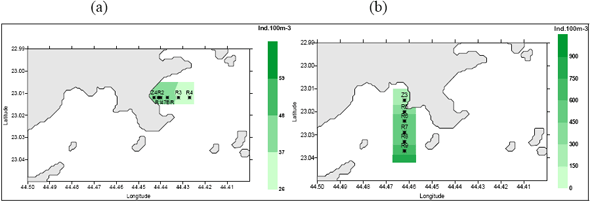
Figura 109 - Distribuição espacial da densidade de ovos de peixes da malha de 500 mm da área de Piraquara de Fora (a) e Itaorna (b).
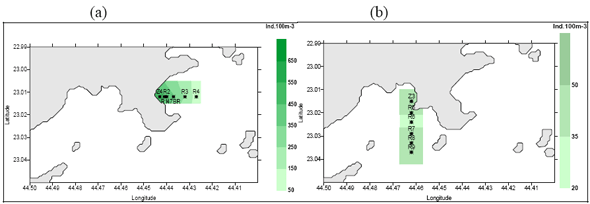
As densidades de larvas, da malha de 330 mm, variaram de 12,60 a 151,79 larvas.100 m-3 e, da malha de 500 mm, de 2,80 a 34,32e larvas.100 m-3(Tabela 94). Apesar das menores densidades das duas malhas da rede bongô estarem em Itaorna, esta área apresentou os valores mais altos de larvas (Figura 110 e Figura 111).
Figura 110 - Distribuição espacial da densidade de larvas de peixes da malha de 330 mm da área de Piraquara de Fora (a) e Itaorna (b).
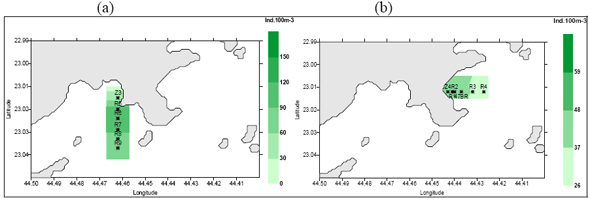
Figura 111 - Distribuição espacial da densidade de larvas de peixes da malha de 500 mm da área de Piraquara de Fora (a) e Itaorna (b) da campanha de maio de 2002.
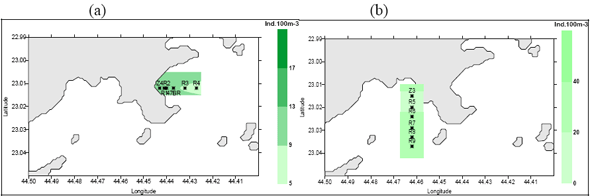
A família Clupeidae foi a mais abundante nas duas malhas da rede bongô, seguidas das famílias Gobiidae e Haemulidae na malha de 330 mm (Tabela 95) e dos Engraulidae e Chloroscombrus chrysurus na malha de 500 mm (Tabela 96). A frequência também foi diferente para as duas malhas da rede bongô. Na malha de 330 µm, as larvas mais frequentes foram de Clupeidae, C. chrysurus e Achirus lineatus, enquanto que, da malha de 500 mm, as mais frequentes foram Engraulidae, Clupeidae, C. chrysurus e Gobiidae. As espécies Eucinostomus sp., Menticirrhus americanus, Abudefduf saxatilis, Parablennius sp., Gobiesox strumosus e Symphurus kyaropterygium foram exclusivas da malha de 330 mm. Prionotus punctatus e Umbrina coroides ocorreram somente na malha de 500 µm.
Tabela 95 - Densidade dos táxons de larvas de peixes (larvas.100 m-3), coletados na malha de 330 µm, nas 12 estações nas áreas de Itaorna e Piraquara de Fora.
Clique aqui para visualizar a tabela 95
Tabela96 - Densidade dos táxons de larvas de peixes (larvas.100 m-3), coletados na malha de 500 µm, nas 12 estações nas áreas de Itaorna e Piraquara de Fora .
Clique aqui para visualizar a tabela 96
A dominância de larvas de Clupeiformes (Engraulidae e Clupeidae) em ambientes costeiros brasileiros é um fato comum e já foi registrado por vários autores em outros ecossistemas (Soares et al., 1991; Katsuragawa et al., 1993; Castro & Bonecker, 1996; Bonecker, 1997; Coelho-Botelho et al., 1999). A dominância da família Gobiidae já foi observada no complexo-estuarino de Paranaguá (Sinque et al., 1982); no sistema estuarino da baía de Vitória (Almeida et al., 2000), entre outros.
Em termos de composição taxônomica, observou-se que ocorreu um maior número de famílias de peixes demersais (10) do que de pelágicos (6), nas duas malhas da rede bongô. Entretanto, em termos de abundância, as larvas pelágicas foram mais abundantes que as demersais (Figura 112).
Figura 112 - Percentual das famílias pelágicas e demersais coletadas com a rede bongô nas áreas de Itaorna e Piraquara de Fora.
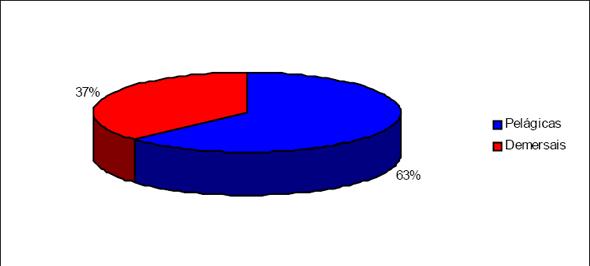
A diferença entre as densidades de peixes pelágicos e o número de famílias de peixes demersais, esta relacionada a abundância dos pequenos pelágicos como as larvas de Clupeidae e Engraulidae. Segundo Katsuragawa et al. (1993), algumas espécies de Engraulidae apresentam um ciclo de vida curto e alta capacidade reprodutiva, permitindo uma rápida expansão populacional.
Microvariação Temporal
Durante a campanha de maio de 2002, em amostragem de microvariação nas áreas de Itaorna (Z3) e Piraquara de Fora (Z4), com a rede bongô, as densidades de ovos de peixes variaram de 100 a 700 ovos.100 m-3 na malha de 330 mm e de 150 a 500 ovos.100 m-3 na malha de 500 mm, com exceção da coleta das 16:50 horas, na maré vazante, onde não foram encontradas larvas de peixes (Tabela 97). A distribuição seguiu o mesmo padrão na área de Itaorna. Contudo, em Piraquara de Fora somente na maré vazante, das 17:55 horas, houve coincidência entre as duas malhas (Figura 113).
Não foram encontrados ovos e larvas de peixes na amostra do ponto fixo em Piraquara de Fora, durante a coleta na maré enchente das 11:30 horas, na malha de 330 mm, devido a grande quantidade de ctenóforos, que dificultou a análise.
Tabela 97 - Intervalos dos horários de coleta, valores de volume, número e densidade de ovos de peixes e de larvas coletados em amostragem de microvariação nas áreas de Itaorna (Z3) e Piraquara de Fora (Z4).
Clique aqui para visualizar a tabela 97
Figura 113 - Variação da densidade de ovos de peixes (ovos.100 m-3) durante a amostragem de microvariação, nas áreas de Itaorna (Z3) e Piraquara de Fora (Z4), nas duas malhas da rede bongô. Maré enchente/alta nos períodos de 11:30-12:05 e 01:03-02:29 e vazante/baixa nos períodos de 16:50-17:55 e 05:02-06:17.
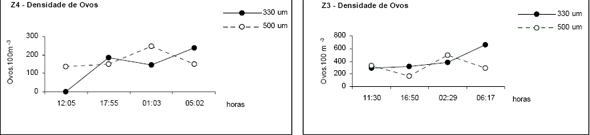
A variação da densidade das larvas de peixes foi semelhante para as duas redes, sendo que a malha de 330 µm apresentou sempre valores mais altos (Figura 114). Matsuura & Nakatani (1980) compararam as densidades de zooplâncton, ovos e larvas de peixes coletados pela rede bongô com malhas de 330 e 500 mm e observaram diferenças significativas entre as malhas e com densidades sempre maiores na malha de 330 µm.
Figura 114 - Variação da densidade de larvas de peixes (larvas.100 m-3) durante a amostragem de microvariação, nas áreas de Itaorna (Z3) e Piraquara de Fora (Z4), nas duas malhas da rede bongô. Maré enchente/alta nos períodos de 11:30-12:05 e 01:03-02:29 e vazante/baixa nos períodos de 16:50-17:55 e 05:02-06:17.
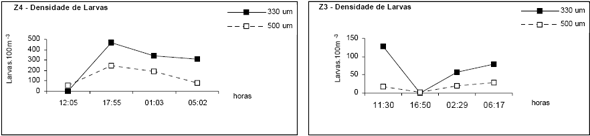
Não foi observada uma relação entre a maré e a variação das densidades de ovos e larvas de peixes. Os engraulídeos dominaram nas duas malhas da rede bongô, seguido dos gobídeos e clupeídeos (Tabela 98 e Tabela 99). As espécies Chloroscombrus chrysurus, Achirus lineatus e Micropogonias furnieri também foram abundantes. Todos os táxons encontrados são comuns em regiões costeiras e semi-fechadas como estuários, baías e lagunas de região tropical.
Tabela 98 - Densidade dos táxons de larvas de peixes (larvas.100 m-3), coletados na malha de 330 µm, durante a microvariação de 24 horas nas áreas de Itaorna (Z3) e Piraquara de Fora (Z4).
Clique aqui para visualizar a tabela 98
Tabela 99 - Densidade dos táxons de larvas de peixes (larvas.100 m-3), coletados na malha de 500 µm, durante a microvariação de 24 horas nas áreas de Itaorna (Z3) e Piraquara de Fora (Z4).
Clique aqui para visualizar a tabela 99
7.4.4.2. Análise das Regiões de Itaorna e Piraquara de Fora - topo
Comparando-se a composição ictioplanctônica entre as áreas de Itaorna e Saco Piraquara de Fora, observou-se uma alternância de dominância para os ovos e as larvas de peixes. Em Itaorna foram observadas as maiores densidades de ovos, tanto no estudo das radiais quanto da microvariação. Para as larvas de peixes, no estudo das radiais, as maiores densidades ocorreram em Itaorna e no estudo da microvariação as maiores densidades foram observadas em Piraquara de Fora.
Apesar desses dados serem de apenas uma campanha, é possível inferir que esta seja uma característica da área.
Sabe-se que alguns peixes desovam durante todo o ano, mas a maioria desova preferencialmente em uma época do ano. A região de estudo está numa área tropical, caracterizada por dois períodos sazonais distintos, um chuvoso e outro seco. Nestas regiões, o período reprodutivo estende-se de outubro a maio e predominam aquelas espécies com desova parcelada (Vazzoler, 1992). Desta forma, é de se esperar que outras épocas do ano favoreçam a reprodução de espécies de peixes que contribuem para o recrutamento local.
No levantamento de dados pretéritos foram encontradas três espécies, seis famílias e duas ordens. Nesta campanha foram coletadas 19 famílias e 20 espécies.
A variedade de larvas de peixes coletadas indica a importância da área como local de desova para estas espécies e reflete a diversidade ambiental que caracteriza este ecossistema.
Os resultados obtidos permitem concluir que a área de Itaorna e Piraquara de Fora pode ser considerada como área de desova e recrutamento para as espécies que aí ocorrem.
O número de táxons registrados e a dominância de larvas pelágicas sobre as demersais são semelhantes a de outros ecossistemas costeiros da costa brasileira, tais como baía de Guanabara e de Sepetiba (RJ), baía de Vitória (ES), estuário do rio Mucuri (BA).
Os táxons encontrados não são endêmicos da área de estudo. As famílias Engraulidae, Clupeidae, Carangidae, Gerreidae, Haemulidae, Sparidae, Sciaenidae e Paralichthyidae tem valor comercial.
7.4.5. Zoobentos - topo
7.4.5.1. Zoobentos de Costão - topo
Em recente levantamento bibliográfico, foi verificado que as pesquisas na orla marítima, na área sob influência da CNAAA, iniciaram na década de 70. Um grupo de pesquisadores liderado pelo Dr Rudolf Barth inventariou a biota e analisou aspectos físico-químicos e geomorfológicos das praias de Itaorna, Mamede e Saco Piraquara de Fora, resultando na montagem de três relatórios (Femar, 1971 a, b e c); desses, apenas dois foram localizados (Femar, 1971 a, e c). Em 1972 alguns pesquisadores, principalmente da USP, estudaram grupos taxonômicos específicos encontrados em áreas pertencentes ao município de Angra dos Reis (Ilha Grande), tendo sido assinalados, na região, equiurídeos (Schaeffer-Novelli, 1972), briozoários (Tommasi et al., 1972), cefalocarídeos (Tommasi et al., 1972), equinodermos (Tommasi, 1974), bivalves (Galvão & Tommasi, 1977; Neme, 1979) e cirripédios (Lacombe, 1977).
Visando fornecer subsídios para o monitoramento da área sob influência da CNAAA-Unidade 1 (Angra 1), após o funcionamento da usina, pesquisadores da UFRJ realizaram, em 1980/1981, um estudo detalhado da biota no Saco Piraquara de Fora e Piraquara de Dentro. Ao término desse minucioso estudo (período pré-operacional), foram indicadas algumas espécies a serem acompanhadas por uma equipe técnica de Furnas Centrais Elétricas S.A. num programa de controle especialmente criado com essa finalidade. Esse acompanhamento resultou em relatórios anuais feitos de 1983 até o presente momento (Programa de Monitoração da Fauna e Flora Marinhas -PMFFM- Zoobenthos “Costão” CNAAA -Unidade 1, 1983; 1986-1988; 1991-1997; 1999-2001). As espécies indicadas na ocasião, para monitoramento, foram Geodia gibberosa, umporífero, Herdmania momus, um ascidiáceo e Tricolia affinis e Diastoma varium, ambos moluscos.
Além dos trabalhos acima mencionados, a bibliografia lista mais uma seqüência de estudos, recentemente desenvolvidos no município de Angra dos Reis, dessa forma contribuindo com um melhor conhecimento da biota das áreas próximas às instalações da atual Eletronuclear (Moraes et al., 1991; Nogueira, 1993; Brasil et al., 1994; Carvalheira et al., 1994; Fonseca et al., 1994; Schmidt et al., 1994; Roiff et al., 1994; Junqueira et al., 1995; Garcia, 1995; Carvalho, 1996; Grillo et al., 1996; Széchy, 1996; Nogueira et al., 1997; Junior et al., 1998 e Széchy et al., 2001).
Como decorrência normal do progresso verificado em países desenvolvidos e naqueles de economia emergente, os ambientes naturais vêm sofrendo, ao longo das últimas décadas, uma série de agressões, de origem antrópica, devido à implantação dos mais diversos tipos de empreendimentos. Em função disso, desde a década de 70 foram criadas, no Brasil, inúmeras leis de proteção ambiental e vêm sendo desenvolvidos em todo o mundo, inclusive no Brasil, vários programas de controle de áreas sob influência desses empreendimentos. A bibliografia especializada aponta uma série de casos de danos causados aos diversos ecossistemas por poluentes de natureza diversa. A poluição decorrente da implantação e, posteriormente, do funcionamento de fábricas, indústrias, usinas e/ou refinarias resulta, em geral, em prejuízos ao ambiente e, conseqüentemente, à própria saúde e bem-estar do homem. O refugo e os efluentes, descartados por tais empreendimentos, na maioria das vezes afetam a qualidade do ar, do solo, dos mananciais hídricos e/ou dos corpos d’água adjacentes.
Ao final da Segunda Guerra, o uso da bomba atômica deixou traumatizada a opinião pública mundial com relação à energia obtida por fissão nuclear. Assim, a implantação de usinas nucleares, para fins pacíficos, passou a estar, sempre, associada a projetos de avaliação ambiental. Esse temor, na verdade, não era infundado; haja visto o terrível episódio ocorrido no final da década de 80 em Chernobil, na Rússia. Entretanto, mesmo descartando a hipótese de risco por contaminação radiativa, o funcionamento de uma usina eletro-nuclear, utilizando água do mar no resfriamento dos condensadores, ocasiona outros tipos de perturbação ambiental, principalmente nas imediações da saída do efluente. Alguns desses distúrbios traduzem-se em elevação da temperatura da água descartada e alterações na sua composição química, durante a passagem pelo sistema de refrigeração, além da contaminação, às vezes em elevadas doses, por biocidas aplicados na água visando evitar a bioincrustação na tubulação.
Constituída por organismos sésseis e vágeis, a biota bentônica é totalmente dependente da qualidade da água onde se encontra. Esta dependência faz com que, em condições adversas, os organismos vágeis se evadam e os sésseis procurem adaptar-se à situação. Mas, quando as condições se tornam críticas, algumas populações acabam sendo extintas. Isto as torna excelentes indicadoras da qualidade da água. O descarte de água quente, como é o caso observado na área estudada, pode ocasionar perturbações que vão, aos poucos, se refletindo na estrutura das comunidades, podendo-se verificar, ao longo do tempo, modificações nos valores de riqueza de espécies, abundância, densidade, biomassa, reprodução, recrutamento, crescimento e, até, o desaparecimento de algumas espécies; em casos extremos até, mesmo, de populações. Mas a elevação da temperatura da água nem sempre causa graves distúrbios. Em países de clima temperado, por exemplo, dependendo da época do ano e das espécies envolvidas, pode-se observar, até mesmo, o crescimento de algumas populações.
Em ambientes marinhos perturbados pelo fator temperatura, um dos pontos mais importantes a considerar é a faixa de “temperatura” ótima característica de cada espécie. As espécies mais sensíveis serão, sempre, aquelas que já se encontram no seu limite máximo de tolerância. Segundo Andrewarta & Birch (1974), animais constantemente expostos a temperaturas extremas, fora do limite de suas faixas favoráveis, podem morrer ou continuar vivos, por períodos ilimitados, sem crescer ou produzir prole. Pelo exposto, a poluição térmica é, sem dúvida, o principal problema existente em áreas onde a biota já se encontra, naturalmente, dentro de seus limites de tolerância térmica, como o que ocorre em países tropicais.
Este diagnóstico reporta os dados obtidos no novo levantamento da área sob influência da CNAAA duas décadas após o primeiro grande inventário realizado na área. As comunidades desta vez analisadas pertencem à chamada “fauna de fital” e o substrato é constituído por algas calcárias articuladas (Amphiroa e/ou Jania). Este fato se deveu a duas razões: primeiro, porque já se supunha encontrar uma diferença ainda que sutil na composição algal da região, comparando com épocas anteriores ao desenvolvimento desta pesquisa; segundo, porque a fauna de fital apresenta uma composição específica típica (Masunari, 1981; 1983; 1989; Masunari & Forneris, 1981), o que, provavelmente, melhor se prestará a um monitoramento. Sempre que possível, os resultados apresentados são comparados com os obtidos em 1980/81.
Os locais estudados foram os costões do Saco Piraquara de Fora, da Praia Brava (Ponta Grande) e da Praia de Fora do Mamede, realizando-se coletas em oito pontos no Saco Piraquara de Fora e dois em Itaorna, como indicado na Figura 115. Para chegar aos locais de coleta foi utilizada uma embarcação e em cada ponto foram anotadas, no local de fundeio da mesma, as coordenadas, a profundidade, a temperatura da água superficial e do fundo e a salinidade. As amostras foram retiradas por mergulhadores na zona correspondente à franja do infralitoral/infralitoral raso, com o emprego de garrafas de ar comprimido.
O material biológico foi obtido por meio de raspagem da rocha, com marreta e talhadeira, sempre que possível destacando o estrato superficial (revestimento de algas calcárias articuladas). Em pontos onde a rocha ofereceu maior resistência à retirada do estrato superficial, foi feita a raspagem total da área limitada pela unidade amostral utilizada, um quadrado de ferro de 25 x 25 cm. Em seguida, o material foi colocado em sacos de tela de náilon, de 500 µm, contendo etiquetas com a identificação do ponto (Pt 2, Pt 9, Pt 26, ...) e o do quadrado (QI, QII, QIII). O número do ponto usado neste trabalho corresponde àquele pintado na encosta do local escolhido para a coleta. Uma vez içadas a bordo, as amostras foram colocadas em baldes com água, do próprio local de coleta, até a chegada ao laboratório de campo. Com o auxílio de um termômetro de mercúrio, foram medidas as temperaturas da água superficial e do fundo e, com um refratômetro. Vista modelo A 366 ATC, a salinidade da água superficial.
No laboratório de campo, foi feita uma triagem grosseira, em grandes grupos, com o objetivo de aplicar ao material o anestésico adequado e agilizar a triagem refinada, feita posteriormente no laboratório do Instituto de Biologia da UFRJ. Após aplicação do anestésico (mentol granulado), o material foi acondicionado em sacos plásticos com etiquetas contendo o nome do projeto, o número do ponto, o local (acidente geográfico mais próximo), o quadrado e a data. Cada amostra foi, então, imersa em solução fixadora de formol a 10% preparada com água do mar.
De cada um dos pontos trabalhados foram colhidas amostras avulsas, para a obtenção de dados complementares qualitativos. Este artifício permite uma análise mais detalhada das comunidades presentes na área, podendo-se coletar algumas espécies comuns que, pelo acaso com que os quadrados foram distribuídos no local amostrado, podem não ter sido contempladas.
No laboratório do Instituto de Biologia da UFRJ (no Departamento de Zoologia), o material foi submetido a triagem fina e em seguida remetido aos especialistas dos principais grupos zoológicos (Cnidaria, Polychaeta, Mollusca, Crustacea e Echinodermata) para a identificação refinada.
A densidade da fauna foi expressa em número de indivíduos pela área do quadrado. A abundância relativa (AR) de cada táxon foi calculada na base de sua proporção em relação à densidade total. A significância dos resultados de densidade total foi avaliada com a aplicação do teste de análise de variância, sendo os dados testados quanto à normalidade e à homocedasticidade (Zar, 1984). Foram calculados os seguintes índices biológicos (Clarke & Warwick, 1994):
- número de táxons = número total de táxons (S) encontrados em cada estaçãodensidade total = número total de indivíduos (N) em cada amostraequitabilidade = índice de Pielou (J)diversidade = índice de Shannon-Wiener (H’)
- dominância = índice de Simpson (SI)
Objetivando visualizar melhor e integradamente a estrutura das comunidades, os resultados dos índices biológicos foram submetidos a análise dos componentes principais. Para a caracterização das associações de espécies, os resultados relacionados à composição e à abundância de espécies foram submetidos a uma análise fatorial de correspondência (Legendre, 1993).
Para o estabelecimento de grupos de estações com composição faunística semelhante, foi aplicado o índice de similaridade de Bray-Curtis (Clarke & Warwick, 1994) aos dados da fauna, transformados pela raiz quadrada. A partir da matriz de similaridade foi feita uma análise de ordenação não métrica multimensional (MDS) dos pontos de coleta (Clarke & Warwick, 1994). Os cálculos foram feitos com o emprego do programa Primer 5.0 for Windows.
Figura 115 - Zôobentos de costão - Postos de coleta
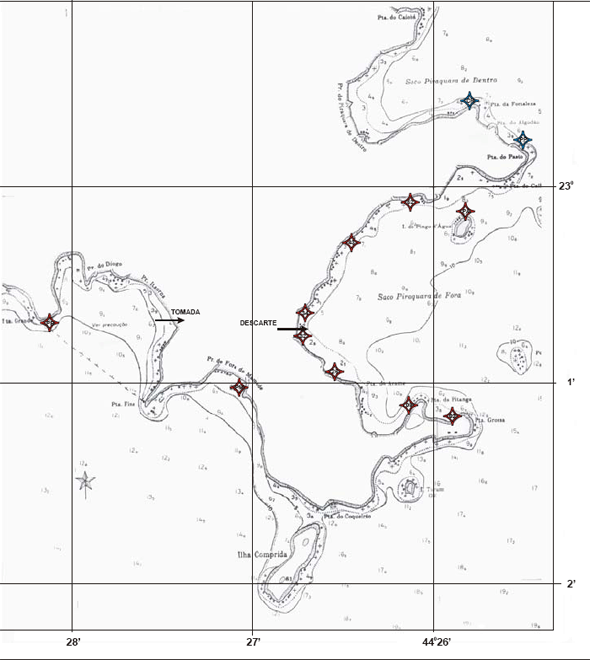
Caracterização dos Zoobentos de Costão
A paisagem submarina dos costões do Saco Piraquara de Fora não é muito diferente da encontrada em todo o litoral do município de Angra dos Reis. O fundo (bentos) é constituído por substrato rochoso, de um modo geral encoberto por algas, algumas delas formando imensos “tapetes”, como é o caso das calcárias articuladas (Amphiroa e Jania, na maioria das vezes), situadas no médio litoral e franja do infralitoral.
Numa zona um pouco mais abaixo (infralitoral raso) podem ser, eventualmente, encontrados “jardins” de Sargassum, sem dúvida outrora bem mais abundantes na região. Essas algas distribuem-se em mosaicos, alternando sua presença com extensos “tapetes” de zoantídeos, cnidários coloniais típicos de coloração amarelo-parda, facilmente identificados pela presença, em sua superfície, de uma camada de muco pegajoso. Neste ambiente, fauna e flora convivem de maneira harmônica, ambos se beneficiando mutuamente.
Marcam presença, ainda, alguns invertebrados facilmente visíveis por mergulhadores amadores, como o tunicado, de coloração escura, da espécie Phallusia nigra e anêmonas do mar, esverdeadas, da espécie Phyllactis flosculifera. O único ponto do costão, de fato, inteiramente descaracterizado, é o de número 32, junto à saída do efluente, onde não mais se avista a fauna e a flora típicas, observando-se a dominância, em escala acentuada, de alguns poliquetos tubícolas cirratulídeos (anelídeos) e siris, crustáceos decápodos carnívoros, predadores vorazes, caracterizando um ambiente inteiramente em desequilibrio.
Neste trabalho foram utilizadas duas estratégias de coleta: uma quantitativa (raspagem dos quadrados) e outra qualitativa (amostras avulsas). A raspagem dos quadrados (QI, QII, QIII) permite quantificar os animais utilizando uma unidade de área conhecida. As amostras avulsas (A) complementam a lista dos táxons presentes na área que, porventura, não tenham sido contemplados nas coletas com os quadrados.
A Tabela 100 contém os dados abióticos anotados durante as coletas feitas nos diversos pontos trabalhados no costão. Nela constam as coordenadas do local de ancoragem do barco, as datas das coletas, a hora e altura das marés, nos dias de coleta, a hora de início das atividades, em cada ponto, as profundidades locais, as temperaturas de superfície e de fundo e as salinidades da água superficial.
Tabela 100 - Dados abióticos tomados nos diversos pontos amostrados no costão do Saco Piraquara de Fora (Pontos 2, 9, 26, 32, 34, 43, 52 e Pingo d’Água-Ilha), na Praia Brava (Ponta Grande) e na Praia do Mamede. X = dados ausentes.
Clique aqui para visualizar a tabela 100
Foi identificado um total de 214 táxons, dos quais 140 (65%) foram comuns às duas estratégias de amostragem utilizadas (Tabela 101).
Tabela 101 - Numero de táxons encontrados neste estudo conforme a estratégia de amostragem utilizada
Número de táxons |
Estratégia de amostragem |
Quadrados |
Avulsas |
Total |
189 |
165 |
Exclusivos |
49 |
25 |
Comuns |
140 |
Considerando a estratégia de amostragem quantitativa (quadrados), foram coletados 28.578 indivíduos representando 14 grandes grupos zoológicos. Desses, o filo Mollusca foi o mais abundante, no total das amostras (43,22%), seguido de Polychaeta (33,57%) e Crustacea (16,41%). Estes três grandes grupos representaram 93,20 % do total de táxons coletados nos quadrados.
A dominância dos grupos variou entre os pontos de coleta. Mollusca foi o grupo dominante nos pontos 2, 26, 34, 43, Pingo d´Água e Praia Brava (Ponta Grande), Annelida nos pontos 9, 32 e Praia de Fora do Mamede e Crustacea no ponto 52 (Tabela 102).
Tabela 102 - Abundância relativa dos grandes grupos zoológicos nos pontos de coleta (%)
Clique aqui para visualizar a tabela 102
Na Tabela 103 encontram-se listadas as 20 principais espécies encontradas, no total das amostras, e suas respectivas abundâncias relativas. As espécies mais abundantes foram Bittium varium e Caecum brasilicum representando, respectivamente, 10,16% e 9,70% no total de táxons coletados. Essas espécies representam gastrópodes que, tipicamente, utilizam macroalgas bênticas como substrato (Rios, 1986; Almeida, 2000). A espécie Cirratulidae morfotipo 1 representou a terceira espécie, em termos numéricos (9,55%) e as espécies Exogone sp. (6,66%), Isognomon sp. (6,36%) e Eunice sp. (6,22%) foram, respectivamente, a quarta, quinta e sexta espécies mais abundantes no total das amostras.
Exogone sp. e Eunice sp. são poliquetos comuns em costões rochosos, já Isognomon sp. é um bivalve exótico, provavelmente introduzido na costa brasileira por água de lastro de navios ou plataformas de petróleo, já tendo sido assinalado em Arraial do Cabo, Rio das Ostras e Macaé (Almeida, observação de campo), onde parece competir por espaço com o mexilhão Perna perna.
Tabela 103 - Abundância relativa das principais espécies no total das amostras
Clique aqui para visualizar a tabela 103
A abundância relativa dessas espécies variou, entre os pontos de coleta, sendo a espécie Bitium varium dominante nos pontos 26, 34, 43 e 52, Caecum brasilicum nos pontos 2 e Praia Brava (Ponta Grande), Exogone sp. no ponto 9; Isognomon sp. em Pingo d´Água e Cirratulidae morfotipo 1 no ponto 32. O ponto localizado na Praia de Fora do Mamede apresentou 3 espécies com os valores de abundância muito próximos, são elas: Exogone sp, Isognomon sp. e Syllinae morfotipo 1 (Tabela 104).
Tabela 104 - Abundância relativa (%) das principais espécies nos pontos de coleta
Clique aqui para visualizar a tabela 104
Os valores dos índices biológicos da fauna, para os pontos de coleta, encontram-se na Tabela 105.
Tabela 105 - Valores dos índices biológicos da fauna nos diversos pontos de coleta
Clique aqui para visualizar a tabela 105
Os valores de densidade total apresentaram acentuada variação, entre os pontos de coleta (Figura 116), sendo os maiores valores de densidade encontrados na Praia Brava (Ponta Grande) e no Ponto 32 e os menores nos pontos 34 e 52. A análise de variância mostrou diferenças significativas nos valores de densidade da fauna.
Figura 116 - Densidade média da fauna total nos pontos de coleta
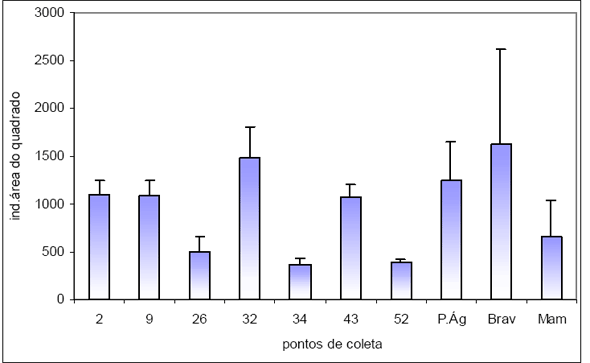
Os maiores valores de diversidade foram encontrados em Pingo d’Água-Ilha e na Praia de Fora do Mamede, sendo o valor mais baixo encontrado no Ponto 32 (Figura 117). Este padrão foi o mesmo para o número de táxons (Figura 118).
Figura 11 7- Diversidade encontrada nos pontos de coleta
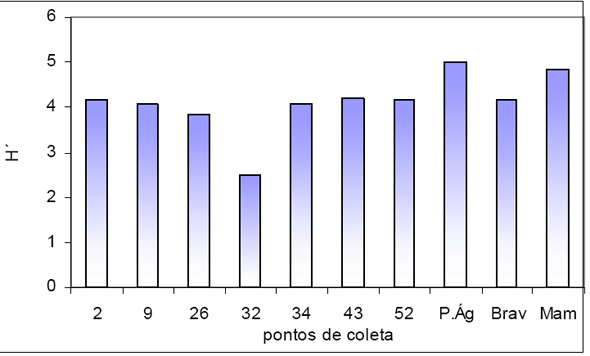
Figura 118 - Numero de táxons presentes nos pontos de coleta
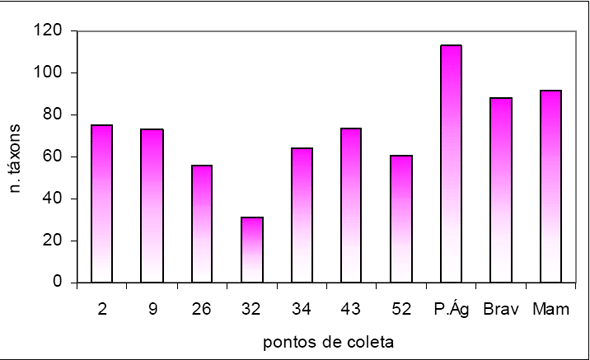
Na sua condição de ilha, próxima ao continente, Pingo d’Água, situada no flanco norte do Saco Piraquara de Fora, apresenta um hidrodinamismo peculiar. A corrente oceânica circula, predominantemente, no sentido do quadrante sudoeste ao quadrante nordeste, renovando, constantemente, a água em seu entorno, mantendo-a límpida e rica em nutrientes, acontecendo o mesmo com as correntes de marés.
A Praia de Fora do Mamede, por sua vez, é diretamente banhada pela água oceânica que incide em seu estirâncio e nos bordos enrocados quase que em ângulo reto. Como praia de alta energia, seu hidrodinamismo acentuado provavelmente interfere um pouco na composição de suas comunidades bênticas, daí os valores de diversidade serem ligeiramente mais baixos.
Já o Ponto 32, situado no interior do Saco Piraquara de Fora, onde, teoricamente, a diversidade deveria ser grande, os valores de densidade espelham, fielmente, o grau de comprometimento local. Trata-se do ponto mais próximo à saída do efluente da usina, onde as características geomorfológicas primárias se encontram inteiramente descaracterizadas em conseqüência de aterros e do enrocamento erguido para a proteção da tubulação.
As equitabilidades ficaram, em geral, em torno de 0,67, sendo o valor mais alto encontrado na Praia de Fora do Mamede e o mais baixo no Ponto 32. A equitabilidade é um bom parâmetro para medir a dominância (Pielou, 1969); quanto mais próximo ela estiver de 1,0 melhor será a distribuição dos indivíduos entre as espécies, ou seja, menor será a dominância.
Ambientes com baixa dominância representam áreas mais estáveis, apresentando comunidades onde os recursos estão sendo bem repartidos entre as espécies, enquanto comunidades com elevada dominância, de uma ou poucas espécies, indicam locais onde estas espécies estão levando vantagem, sobre as demais, com relação aos recursos disponíveis. Esta segunda alternativa ocorre, em geral, em ambientes estressados. Esse padrão pode ser observado, também, pelo índice de dominância de Simpson que, com exceção do Ponto 32, mostrou valores inferiores a 0,15 (Tabela 105).
A análise em componentes principais (PCA), realizada com os valores dos índices biológicos da fauna, possibilitou uma análise conjunta dos pontos de coleta (Figura 119). Os que apresentaram os maiores valores de densidade foram o 32 e Praia Brava (Brav), enquanto Pingo d’Água (PÁg) e Praia do Mamede (Mam), apresentando as diversidades e as equitabilidades mais elevadas, mostraram as comunidades melhor estruturadas; tais comunidades, em geral, têm a capacidade de suportar melhor as ações causadoras de estresse. Os demais pontos apresentaram estruturas de comunidades semelhantes.
Figura 119 - Análise em componentes principais (PCA) aplicada sobre os valores dos índices biológicos.
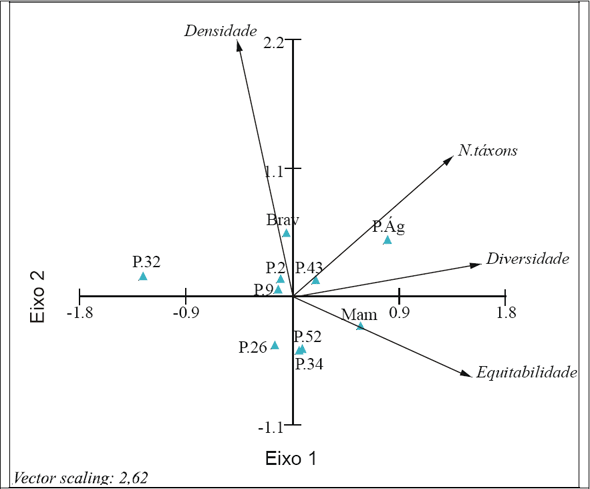
A fim de caracterizar as associações de espécies, foi realizada uma análise fatorial de correspondência considerando, nesta caracterização, apenas as espécies com abundâncias relativas superiores a 0,70. Essas espécies somaram 75% do total de organismos coletados. A análise fatorial aplicada a essas espécies evidenciou uma composição distinta entre o Ponto 32 e os demais, tendo o fato sido causado pelas abundâncias de Polydora sp e Cirratulus sp. (Figura 120).
Como o Ponto 32 apresentou características distintas dos demais, podendo ter interferido nas caracterizações, uma nova análise de correspondência foi efetuada excluindo o mesmo. O resultado desta nova análise pode ser visto na Figura 121.
Figura 120 - Análise fatorial de correspondência realizada a partir das abundâncias das principais espécies
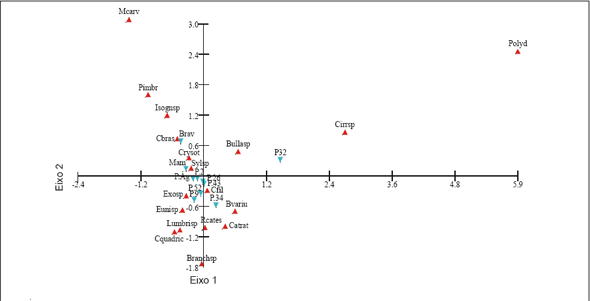
Figura 121 - Análise fatorial de correspondência realizada a partir das abundâncias das principais espécies, excluindo o Ponto 32.
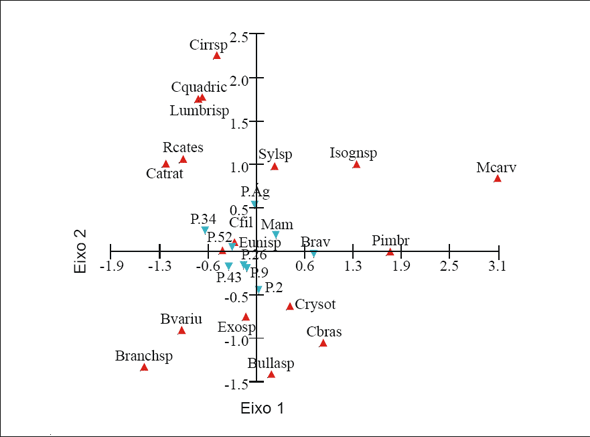
Os eixos 1 e 2 explicaram, aproximadamente, 60% da variação nas abundâncias das espécies, nos diferentes pontos de coleta (Figura 121). Praia Brava (Ponta Grande) foi o ponto que apresentou uma associação de espécies, distinta das demais, constituída por Modiolus carvalhoi, Pinctada imbricata e Isognomon sp.
Cabe aqui, uma vez mais, ressaltar que o bivalve Isognomon sp é uma espécie exótica, provavelmente introduzida por águas de lastro de navios ou plataformas. Esta espécie compete com as espécies nativas e a forma com que vem se disseminando, por todo o litoral, sugere que poderá vir a acarretar, em médio e longo prazo, profundas alterações na estrutura e composição das comunidades bênticas dos substratos consolidado e do fital.
Ao longo do eixo 2 foram evidenciadas duas outras associações, uma em Praia Brava (Ponta Grande), constituída por Bulla sp, Branchiomma sp, Caecum brasilicum, Bittium varium, Exogone sp e Caecum rysotitum e outra em Pingo d´Água-Ilha, formada por Modiolus carvalhoi, Syllis sp, Isognomon sp, Cerithium atratum, Rissoina catesbyana, Lumbrineris sp, Corophium quadriceps e Cirratulus sp.
A análise de ordenação não métrica multimensional (MDS) evidenciou a formação de 5 grupos de pontos de coleta. O primeiro grupo é formado pelos pontos de coleta 34, 32 e 26. O segundo é formado pelos pontos 9 e 43, o terceiro pelos pontos 2 e 52 e o quarto é representado somente pelo ponto localizado em Pingo d´Água-Ilha. O quinto grupo foi formado pelos pontos localizados fora do Saco Piraquara de Fora, ou seja, Praia Brava (Ponta Grande) e Praia de Fora do Mamede (Figura 122).
Figura 122 - Análise multidimensional (MDS) baseada na densidade dos táxons nos pontos de coleta
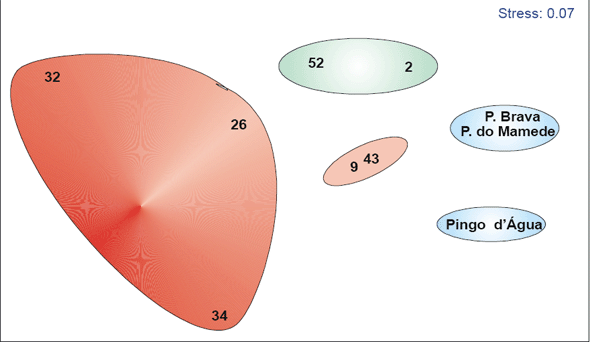
Utilizando os grupos de pontos de coleta observados, a partir da análise de ordenação multidimensional (MDS), podemos definir que a estrutura das comunidades estudadas responde a um “gradiente ambiental” provavelmente provocado por influência do efluente. A distribuição espacial dos pontos de coleta, em função desse gradiente, pode ser visualizada na Figura 123. Assim, temos o primeiro grupo de pontos (32, 34 e 26) definido como aquele mais próximo à saída do efluente (A). O segundo (pontos 43 e 9) e o terceiro (pontos 2 e 52) grupos representam áreas adjacentes à saída do efluente (B e C). O quarto grupo (D) é representado pelo ponto mais afastado da saída do efluente (Pingo d’Água-Ilha), mas ainda localizado no Saco Piraquara de Fora. O quinto grupo (E), representado pelos pontos Praia Brava e Praia do Mamede, fica localizado em áreas fora do Saco Piraquara de Fora (Figura 123).
Figura 123 - Representação, baseada na análise de ordenação, dos pontos de coleta em função do gradiente ambiental causado pelo efluente situado no Saco Piraquara de Fora
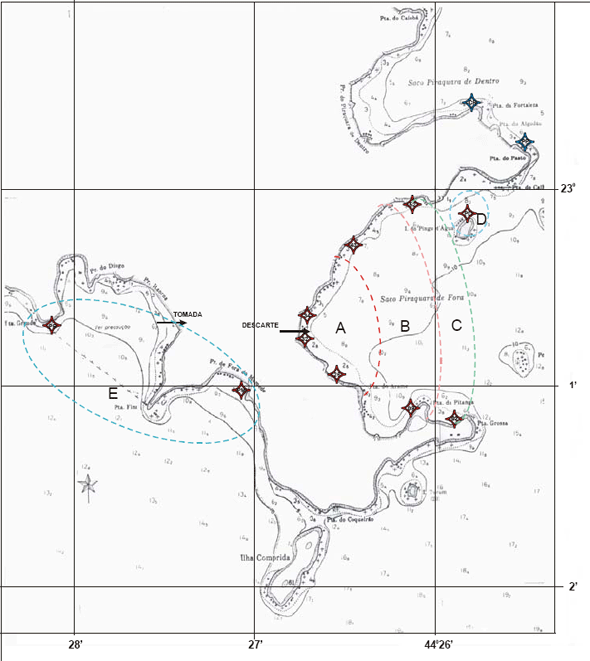
Citando palavras atribuídas a Drach (1951), l`Hardy (1962) já comentava que a riqueza de espécies encontrada em costões rochosos marinhos devia-se, essencialmente, à presença do revestimento algal, refúgio apreciado por inúmeros animais vágeis, contra a predação, e oferta de substrato complementar à fixação de espécies sésseis.
A distribuição da epifauna nessas algas, por sua vez, dependia de alguns fatores físicos, principalmente do nível de oscilação das marés, do fluxo d’água, da turbulência e da deposição de sedimento no substrato algal. Sob o ponto de vista ecológico, a epibiose oferece alguns problemas tais como variações sazonais das populações epífitas, ação de fatores ecológicos sobre estas populações e modalidade de renovação das espécies fixas, entre outros. Diversos autores já se dedicaram a estudos de populações epífitas em algas.
Na Europa, por exemplo, alguns dos substratos algais já estudados pertencem aos gêneros Laminaria (Colman, 1939; l’Hardy, 1962), Fucus (Colman, 1939; Hagerman, 1966; Oswald & Seed, 1986) e Ascophyllum (Colman, 1939), entre outros típicos de regiões temperadas. Nas regiões tropicais são mais importantes as algas vermelhas, particularmente as calcárias (Chapman, 1946 apud Masunari & Forneris, 1981), embora poucos pesquisadores tenham, até o momento, se dedicado ao seu estudo. No Brasil pode-se mencionar Gouveia & Leite (1980 apud Masunari, 1983), trabalhando com Halimeda e Masunari (1981; 1982; 1983) com Amphiroa, algas calcárias articuladas.
Segundo Masunari (1989), o termo fital foi proposto por Remane, em 1933, para designar um habitat marinho dominado por macrófitas, onde coexistem animais e plantas epífitas. Levando em consideração as características próprias e a capacidade de manutenção dessas características, autores como Masunari & Forneris (1981) promoveram o fital a um ecossistema distinto. Até o final da década de 60, estudos sobre o fital, no Brasil, levavam em consideração apenas a fauna vágil ou a fauna séssil. Estudos envolvendo os organismos de forma integrada tiveram início com Masunari (1981; 1982; 1983) utilizando Amphiroa beauvoisii e A. fragilissima do litoral de São Paulo, onde essas calcárias articuladas ocupam, preferencialmente, o médio e o infralitoral raso sob a forma de extensos tapetes de 0,70 a 2,50m de largura, com até 40mm de altura.
Analisando 99 amostras do fital Amphiroa beauvoisii coletadas no médio litoral e infralitoral superior, no inverno e no verão, Masunari (1981) identificou 172 táxons (entre organismos vágeis e sésseis), contando 93.916 espécimes, enquanto neste estudo, em apenas 30 amostras quantitativas obtidas no infralitoral superior, em dez pontos trabalhados em uma única campanha foram identificados 191 táxons de organismos vágeis e contados 28.578 indivíduos, a maioria sobre A. fragilissima. Em sua tese Masunari (1981) verificou que os grupos zoológicos mais representativos foram Polychaeta, Mollusca e Crustacea, Polychaeta e Mollusca apresentando o maior número de espécies e Crustacea o maior número de indivíduos.
Durante os levantamentos (2002), os grupos que se destacaram foram Mollusca, constituindo o grupo mais rico em táxons (64, contra 60 em 1980/81 e 28 em Masunari, 1983), seguidos de Crustacea (63, contra 68 em 1980/81 e 39 em Masunari, 1983) e Annelida Polychaeta (37, contra 72 em 1980/81 e 42 em Masunari, 1983). Foram os Mollusca, também, que apresentaram o maior número de indivíduos, principalmente as espécies Bittium varium (2904 ind.)e Caecum brasilicum (2773 ind.). Pelo exposto, verifica-se que a fauna de poliquetos apresentava quase o dobro de táxons na década de 80 (72 táxons em 1980/81, contra 37 de agora). Com relação à fauna de moluscos, praticamente não foi notada variação da década de 80 para os dias atuais (64 táxons em 1980/81, contra 61 de agora), podendo-se dizer mesmo com relação à fauna de crustáceos (68 táxons em 1980/81, contra 63 de agora).
Existem várias explicações plausíveis para o fato; a primeira é que, em 1980/81, a faixa do litoral trabalhada foi o ecótono médio litoral, interface entre a terra e o mar, enquanto agora a faixa estudada se deslocou para a franja do infralitoral/infralitoral raso; a segunda é que em 1980/81 as coletas foram realizadas mensalmente, enquanto agora elas foram feitas apenas em uma estação (outono); a terceira é que Amphiroa talvez não seja o substrato mais apreciado pelos poliquetos. Outras cogitações poderiam, ainda, ser feitas. Quanto aos moluscos, como o grupo mais bem representado na campanha de 2002, não só no número de táxons como em abundância, eles, provavelmente, tiveram uma chance maior de serem contemplados na unidade amostral utilizada do que os demais grupos.
Segundo a bibliografia especializada não é incomum, em ambientes naturais, a mortalidade de determinadas populações que constituem suas biotas típicas em função de extremos físicos eventualmente ocorridos. Na maioria das vezes os principais fatores ambientais (temperatura, salinidade, luminosidade e teor de oxigênio dissolvido, entre outros) encontram-se intimamente interligados, daí a dificuldade em se analisar os efeitos de um fator, isoladamente, sobre a biota numa dada área de estudo.
Na década de 70, Andrewartha & Birch (1974) afirmavam que a faixa de temperatura favorável a uma dada espécie, em particular, quase sempre coincidia com as temperaturas prevalecentes nos locais onde ela normalmente era encontrada. Na verdade, até o momento poucos foram os estudos feitos com invertebrados marinhos, típicos de regiões entre marés e do infralitoral raso, abordando temas como preferência e/ou níveis de tolerância a temperaturas baixas e/ou altas.
Mas alguns experimentos feitos com invertebrados mostraram uma surpreendente preferência dos animais, objetos de estudo, por uma faixa de temperatura maior do que a encontrada nos seus habitats naturais (Meadows & Ruagh, 1981). Utilizando temperaturas variando de 00C a 350C, estes autores verificaram que o anfípodo Corophium volutator (Pallas) mostrou, em laboratório, preferência pela faixa compreendida entre 150C e 200C, embora em seu habitat natural a temperatura média anual oscilasse entre 00C e 120C. Teoricamente não existe uma razão ecológica óbvia para explicar este comportamento; supõem, os autores, que ele se deva à grande flutuação da temperatura a que costuma estar exposta a zona entre marés. O experimento citado, no entanto, foi realizado com um animal típico de zona temperada, aonde, em condições normais, a temperatura da água do mar não chega a extremos críticos.
Moore (1940b, apud Andrewartha & Birch, 1974) listou as faixas favoráveis de 41 grupos animais, entre crustáceos, equinodermos, moluscos, tunicados e peixes, verificando que, para 37 delas (91%), a extensão dessa faixa era de cerca de 160C ou menos; para 17 (42%) ela era menor que 140C, ficando duas espécies de peixes restritas a uma faixa de cerca de 80C. Pelo exposto, nota-se que a faixa de temperatura ideal varia não só com o tipo de ambiente estudado (fatores extrínsecos) como, também, com o grupo zoológico considerado (fatores intrínsecos). Em experimentos feitos com cracas, Southward (1958) concluiu que existem, ainda, diferentes faixas de temperatura toleráveis, dependendo do estado fisiológico considerado. Alguns animais são capazes de suportar temperaturas extremas e sobreviver durante um determinado tempo, só que isto acontece com o prejuízo de algumas de suas funções vitais: a reprodução, por exemplo. Mas todos os exemplos acima citados referem-se a experimentos feitos em laboratório, quando, na natureza, as coisas podem não funcionar da mesma maneira.
As curvas traçadas na Figura 123 mostram que, teoricamente, a riqueza de espécies aumenta conforme se afasta, gradativamente, do fundo da enseada para seus bordos externos (A=>B=>C), provavelmente devido ao tipo de distúrbio observado na área sob influência direta do descarte do efluente. Ao mergulhar para fazer as coletas, no Ponto 32, a equipe de campo constatou, na água, um odor característico de cloro, além de uma diferença, marcante, nas temperaturas de superfície comparadas com as de fundo (ver Tabela 100). Em alguns pontos trabalhados foi assinalada uma diferença de até 5ºC, o que não é um fato corriqueiro, em ambientes marinhos naturais de zonas tropicais, a não ser quando ocorre ressurgência. A fauna, provavelmente, se ressente das condições atualmente existentes no Saco Piraquara de Fora, podendo, as comunidades presentes, já não serem as mesmas existentes em épocas anteriores ao funcionamento da usina. As observações feitas neste estudo mostram que o comportamento verificado com relação às populações vágeis também se repete nas populações sésseis.
Dessa maneira, a área estudada encontra-se, de fato, alterada em função da influência do efluente liberado pela CNAAA. A alteração verificada ao longo do Saco Piraquara de Fora se traduz em um “gradiente de melhora”, já esperado, no sentido do fundo da enseada para seus bordos externos, onde os pontos 32 (lado direito) e 34 (lado esquerdo), adjacentes à saída do efluente, se mostram como os mais comprometidos neste momento. A Ilha de Pingo d’Água ainda é o local que apresenta as melhores condições.
Em termos comparativos, as praias de Fora do Mamede e Brava (Ponta Grande) apresentaram condições compatíveis com o esperado para praias oceânicas.
7.4.5.2. Zoobentos de Sedimento - topo
Considerando, especificamente, a área sob influência da Central Nuclear Almirante Álvaro Alberto (CNAAA), verifica-se que as informações gerais sobre a fauna de sedimento de fundo obtidas anteriormente foram baseadas, principalmente, em relatórios técnicos elaborados (Femar, 1971; UFRJ, 1981, Furnas, 1983, 1989, 1990, 1991, 1992, 1993, 1994, 1995, 1996, 1997; Eletronuclear, 1998, 1999, 2000, 2002).
Assim, a geração de informações primárias, ao longo de um período de tempo extenso, representa o mais importante acervo sobre o zoobentos desta região e, até mesmo, de toda a Baía da Ribeira. Somando-se a esses estudos, existem poucos dados publicados em artigos científicos.
O de Manso & Absalão (1988), baseado nos dados obtidos em 1980 (UFRJ, 1981), tornou-se o único trabalho feito com uma comunidade bentônica de fundo na área sob influência do empreendimento. Estes autores estudaram a distribuição dos Ophiuroidea nos Sacos Piraquara de Dentro e de Fora, listando 10 espécies desta classe, dessa forma contribuindo com informações para o monitoramento dos impactos relacionados ao efluente da usina. Por este motivo, optou-se em dar ênfase à comparação dos dados obtidos em 2002 com a série temporal de 1981 a 2000.
Devem ser levadas em consideração as diferenças metodológicas, por exemplo, diferentes amostradores e abrangência da comunidade, e/ou a falta de informações, p. ex., sobre a área do amostrador utilizado de 1982 a 2000, o que limitou a comparação dos dados.
Os pontos de coleta foram estabelecidos com base nos estudos anteriores, que concentraram a amostragem nas áreas controle (Itaorna) e de impacto (Piraquara de Fora), tendo sido estabelecidas 10 pontos, sendo oito no Saco Piraquara de Fora e dois em Itaorna (Tabela 106 e Figura 124), procurando posicioná-los o mais próximo possível das estações utilizadas no monitoramento.
Tabela 106 - Zôobentos de sedimento - Dados dos pontos de coleta
Clique aqui para visualizar a tabela 106
O posicionamento foi feito com a utilização de GPS Garmin, sendo a profundidade determinada por ecosonda portátil da Speedtech Instruments. Dificuldades ocorridas no campo levaram a alterações no posicionamento, tendo o caso mais típico ocorrido na estação 7 (correspondente à estação Z4), onde, devido à forte correnteza provocada pelo efluente das usinas Angra 1 e 2, foi necessário deslocar o local de coleta, no sentido Norte, o que não chegou a alterar as coordenadas porque o deslocamento foi pequeno.
O equipamento de coleta utilizado foi um busca-fundo do tipo van Veen, com uma área de amostragem de 0,06m2. Em cada estação foram coletadas três réplicas do sedimento de fundo e de cada réplica foram retirados cerca de 50 gramas para análise da composição granulométrica. As amostras biológicas foram lavadas com o emprego de sacos de náilon com abertura de malha de 0,5mm. O sedimento retido foi colocado em um recipiente de plástico, fixado em formol a 10% com água do mar, identificado com rótulos contendo as informações da estação de coleta. As amostras assim preservadas foram embaladas e enviadas ao laboratório para análise.
Em laboratório, sedimento retido nas peneiras de 0,5 mm foi triado sob magnificação, com a utilização de microscópio estereoscópico. Os animais foram separados segundo seus filos, acondicionados em frascos rotulados e enviados para a identificação taxonômica. Na identificação procurou-se chegar ao nível taxonômico mais baixo e, quando isto não foi possível, o material foi identificado em categorias taxonômicas altas (filo ou classe).
Para a análise granulométrica, as amostras foram secas em estufa a 100°C por um período de 24 horas e, a seguir, peneiradas de acordo com a metodologia descrita por Buchanan (1984). Nas amostras em que os teores de finos foram superiores a 5%, a fração de finos foi obtida por pipetagem, em tubos de sedimentação, com adição de solução desfloculante de carbonato de sódio e hexametafosfato de sódio. As velocidades de decantação foram calculadas para pesos específicos de 2,65 g.cm-3 (Buchanan, 1984). A partir dos resultados obtidos, foram calculados os parâmetros granulométricos, quais sejam, tamanho médio do grão (Mz), grau de seleção (DPG), assimetria (Ski) e curtose normalizada (K´n) (Folk & Ward, 1957).
A significância dos resultados de tamanho médio dos grãos foi avaliada mediante a aplicação do teste de análise de variância, testando-se os dados quanto à normalidade e à homocedasticidade (Zar, 1984). A partir do resultado da análise de variância, realizou-se o teste de contrastes de Tukey (Zar, 1984). Para o estabelecimento de grupos de estações com características granulométricas semelhantes foi aplicado o índice de distância Euclidiana (Clarke & Warwick, 1994) aos valores dos parâmetros granulométricos utilizados.
Figura 124 -Zôobentos de sedimentos - Pontos de coleta
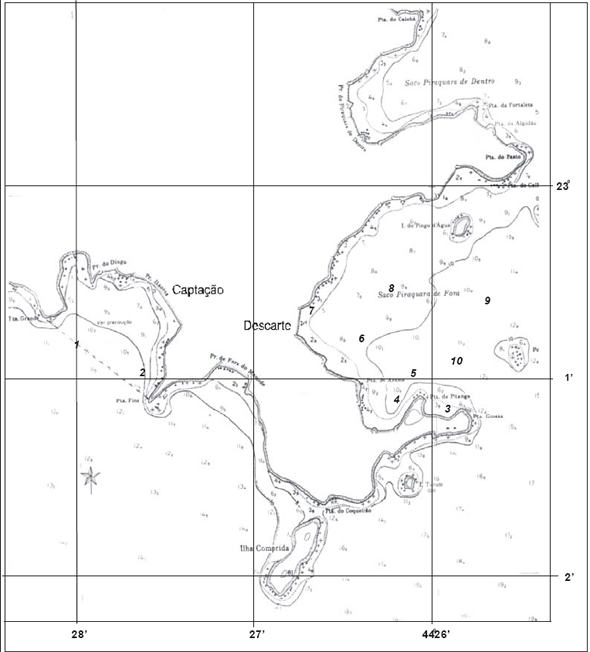
A densidade da macrofauna foi expressa em número de indivíduos por metro quadrado. A abundância relativa (AR) de cada táxon foi calculada na base de sua proporção em relação à densidade total.
A significância dos resultados de densidade da macrofauna total foi avaliada mediante a aplicação do teste de análise de variância, testando-se os dados foram quanto à normalidade e à homocedasticidade (Zar, 1984).
Foram calculados os seguintes índices biológicos para a fauna (Clarke & Warwick, 1994):
-
número de táxons = número total de táxons (S) encontrados em cada estação;
-
densidade total = número total de indivíduos (N) em cada amostra;
-
equitabilidade = índice de Pielou (J);
-
diversidade = índice de Shannon-Wiener (H’);
-
dominância = índice de Simpson (SI).
Para o estabelecimento de grupos de estações com composição faunística semelhante, foi aplicado o índice de similaridade de Bray-Curtis (Clarke & Warwick, 1994) aos dados da fauna, transformados pela raiz quadrada. Para a significância dos resultados da análise de similaridade foi aplicado o teste Anosim (Clarke & Warwick, 1994). A matriz de similaridade de Bray-Curtis foi utilizada também para a análise de ordenação não métrica multimensional (MDS) das estações de coleta (Clarke & Warwick, 1994). Os cálculos foram efetuados com o emprego do programa Primer 5.0 for Windows.
Dufrêne & Legendre (1997) descreveram um índice simples para indicar espécies e/ou associações de espécies que caracterizam grupos de amostras/estações. Para o cálculo desse índice, foram utilizados os valores de densidade e freqüência de cada gênero, e os grupos de amostras foram determinados a partir dos padrões observados na análise de similaridade. A significância dos valores do índice foi testada pelo método de aleatorização descrito por Connor & Simberloff, também conhecido como teste de Monte Carlo (Paes & Blinder, 1995), tendo sido realizadas 1.000 iterações.
Para uma avaliação da relação entre a fauna e o sedimento foi feita uma análise de correlação de Spearman entre a matriz de similaridade da fauna e a matriz de distância dos parâmetros granulométricos (Clarke & Warwick, 1994), sendo os cálculos realizados com o emprego do programa Primer 5.0 for Windows.
As abreviaturas utilizadas nas tabelas e figuras são apresentadas abaixo:
| AM |
Areia média |
MBS |
Muito bem selecionado |
| AMN |
Assimetria muito negativa |
ML |
Muito leptocúrtica |
| AMP |
Assimetria muito positiva |
MMS |
Muito mal selecionado |
| NA |
Assimetria negativa |
MoBS |
Moderadamente bem selecionado |
| AP |
Assimetria positiva |
MPl |
Muito platicúrtica |
| BS |
Bem selecionado |
MS |
Mal selecionado |
| DPG |
Desvio padrão gráfico |
MoS |
Moderadamente selecionado |
| Chipap |
Chione paphia |
Mz (f) |
Tamanho médio do grão (phi) |
| Codcos |
Codakia costata |
ns |
não significativo |
| Corcar |
Corbula caribaea |
Nucsem |
Nucula semiornata |
| Dengou |
Dentalium gouldii |
p |
nível de significância |
| Densp |
Dentalium sp |
Pl |
Platicúrtica |
| Edwsp |
Edwardsia sp |
Pitsp |
Pitar sp. |
| g.l. |
graus de liberdade |
s |
Significativo |
| K´n |
Curtose normalizada |
Ski |
Assimetria |
| L |
Leptocúrtica |
Sim |
Simétrico |
| Meso |
Mesocúrtica |
Telsan |
Tellina sandix |
Granulometria
Na área de estudo predominaram os sedimentos finos e muito mal selecionados a moderadamente selecionados (Tabela 107).
Tabela 107 - Parâmetros granulométricos (valores médios) nas estações de coleta.
| Estações |
MZ |
DP |
Ski |
K’n |
| Valor |
Classificação |
Valor |
Classificação |
Valor |
Classificação |
Valor |
Classificação |
| 1 |
3.63 |
AMF |
0.42 |
BS |
0.01 |
Sim |
0.58 |
L |
| 2 |
1.48 |
AM |
0.83 |
MoS |
-0.05 |
Sim |
0.61 |
ML |
| 3 |
1.38 |
AM |
0.90 |
MoS |
-0.25 |
NA |
0.53 |
L |
| 4 |
6.92 |
S |
2.5 |
MMS |
0.22 |
AP |
0.41 |
Pl |
| 5 |
6.49 |
S |
2.67 |
MMS |
0.41 |
AMP |
0.37 |
MPl |
| 6 |
6.96 |
S |
2.51 |
MMS |
0.21 |
AP |
0.4 |
Pl |
| 7 |
3.19 |
AMF |
1.04 |
MoS |
0.03 |
Sim |
0.6 |
L |
| 8 |
6.02 |
S |
2.32 |
MMS |
0.48 |
AMP |
0.45 |
Pl |
| 9 |
3.68 |
AMF |
0.91 |
MoS |
0.04 |
Sim |
0.65 |
ML |
| 10 |
3.67 |
AMF |
0.67 |
MoBS |
0.12 |
AP |
0.73 |
ML |
A análise de variância, realizada com os valores de tamanho médio do grão, mostrou diferenças significativas entre as estações, sendo observada, pelo teste de contrastes de Tukey, a formação dos seguintes grupos de estações: estações 2 e 3; estações 1, 7, 9 e 10 e estações 4, 5, 6 e 8.
Os resultados observados nas análises de significância foram corroborados pela análise de grupamento, que evidenciou a formação desses mesmos grupos de estações (Figura 125).
Figura 125 - Dendrograma resultante da aplicação do índice de Distância Euclidiana aos valores obtidos para os parâmetros granulométricos.
Os valores numéricos dos resultados granulométricos obtidos em 1980/81 não foram discriminados (UFRJ, 1981), mas as texturas granulométricas encontradas corresponderam às indicadas na Tabela 107. O sedimento bem selecionado, encontrado na estação 1, indicou que o hidrodinamismo neste local é mais intenso do que nas demais estações de coleta.
Fauna
Na Tabela 108 encontram-se discriminados os valores dos índices biológicos da fauna para as estações de coleta.
Tabela 108 - Indices biológicos da fauna nas estações de coleta.
| Estações |
N. táxons |
Densidade |
Equitabilidade |
Diversidade |
Dominância |
| 1 |
29 |
1144.44 |
0.64 |
3.15 |
0.22 |
| 2 |
64 |
4088.89 |
0.80 |
4.78 |
0.06 |
| 3 |
39 |
5661.11 |
0.67 |
3.52 |
0.17 |
| 4 |
29 |
1116.67 |
0.76 |
3.71 |
0.13 |
| 5 |
32 |
1050.00 |
0.70 |
3.52 |
0.18 |
| 6 |
18 |
400.00 |
0.75 |
3.11 |
0.18 |
| 7 |
40 |
1688.89 |
0.82 |
4.36 |
0.07 |
| 8 |
25 |
1233.33 |
0.66 |
3.08 |
0.21 |
| 9 |
69 |
2500.00 |
0.81 |
4.94 |
0.05 |
| 10 |
56 |
2627.78 |
0.79 |
4.57 |
0.08 |
Em valores absolutos foram coletados 3872 indivíduos que, de acordo com a área do amostrador, correspondem a uma densidade média 2151 org/m2, distribuídos em 144 táxons. Esses táxons representam 12 grandes grupos zoológicos, dos quais o filo Mollusca foi o mais abundante em todas as estações de coleta (Tabela 109). A dominância absoluta dos Mollusca na área de estudo pode ser visualizada na Figura 126.
Tabela 109 - Abundância relativa (%) média dos grandes grupos nas estações
| Grupos |
Estações |
| 1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
| Cnidaria |
0.49 |
0.68 |
0.00 |
0.00 |
0.00 |
1.39 |
1.32 |
0.45 |
2.22 |
4.23 |
| Platyhelminthes |
0.00 |
0.27 |
0.10 |
0.00 |
0.53 |
0.00 |
0.33 |
0.00 |
0.00 |
0.85 |
| Nemertinea |
0.00 |
0.14 |
0.10 |
0.00 |
0.00 |
1.39 |
0.00 |
0.90 |
0.89 |
1.48 |
| Sipuncula |
0.00 |
0.14 |
0.00 |
0.00 |
0.00 |
0.00 |
0.00 |
0.00 |
1.56 |
0.63 |
| Echiura |
0.00 |
0.00 |
0.00 |
0.50 |
0.00 |
0.00 |
0.00 |
0.00 |
0.44 |
0.00 |
| Annelida |
9.71 |
30.84 |
10.21 |
4.48 |
16.93 |
13.89 |
28.29 |
18.02 |
32.44 |
15.64 |
| Mollusca |
67.96 |
52.58 |
76.94 |
93.03 |
76.19 |
77.78 |
53.62 |
77.93 |
56.00 |
67.23 |
| Crustacea |
21.84 |
10.33 |
10.60 |
1.49 |
3.17 |
4.17 |
16.45 |
2.70 |
5.78 |
8.88 |
| Echinodermata |
0.00 |
0.14 |
0.20 |
0.00 |
1.06 |
1.39 |
0.00 |
0.00 |
0.44 |
1.06 |
| Hemichordata |
0.00 |
0.00 |
0.00 |
0.50 |
2.12 |
0.00 |
0.00 |
0.00 |
0.00 |
0.00 |
| Chordata |
0.00 |
4.89 |
1.86 |
0.00 |
0.00 |
0.00 |
0.00 |
0.00 |
0.22 |
0.00 |
Figura 126 - Abundância relativa dos grandes grupos nas estações de coleta.
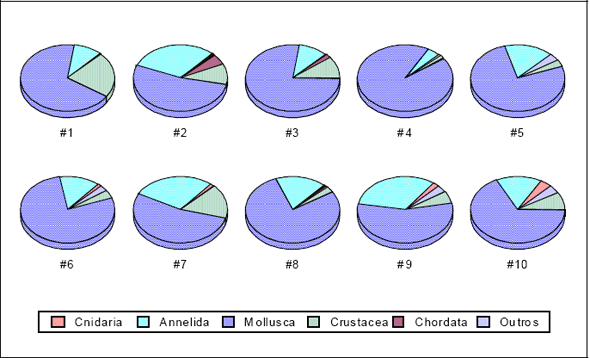
Como conseqüência desta dominância, das 26 espécies que correspondem a cerca de 80% da fauna, 14 são Mollusca (Tabela 110). Semele nuculoides e Codakia costata apresentaram-se como as de maior abundância relativa no total das amostras, perfazendo, respectivamente, 13,97% e 11,21% do total de indivíduos coletados.
Tabela 110 - Abundância relativa (%) das principais espécies no total das amostras.
Clique aqui para visualizar a tabela 110
A espécie Finella dubia representou a terceira espécie numericamente mais importante (6,48%), e as espécies Acteocina bidentata (5,99%) e Nucula semiornata (5,04%) foram, respectivamente, a quarta e quinta espécies mais abundantes no total das amostras. As espécies Acteocina bullata, Transenella stimpsoni, Antallis sp., Caecum brasilicum, Tellina sandix, Crenella divaricata, Tellina sp., Caecum striatum Abra lioica também se mostraram importantes na estrutura da comunidades.
O segundo grupo em importância foi Annelida e as espécies mais abundantes foram: Polygordius sp., Hemipodus sp., Prionospio sp.1, Magelona sp., Haploscoloplos sp. e Magelona cincta.
O terceiro grupo foi Crustacea, sendo as espécies de Amphipoda Ampelisciphotis podophtalma, Tiborunella viscana e Ampelisca cristata as mais abundantes desse grupo.
Cnidaria foi o quarto grupo em importância com uma única espécie abundante: Edwardsia sp.. Ainda com relação à abundância, nos demais grupos não se destacaram espécies importantes.
Deve ser ressaltado que das 26 espécies mais abundantes, 9 são responsáveis por cerca 60% da fauna e pertencem ao Filo Mollusca (Tabela 110 e Figura 127).
Figura 127 - Abundância relativa das principais espécies no total das amostras.
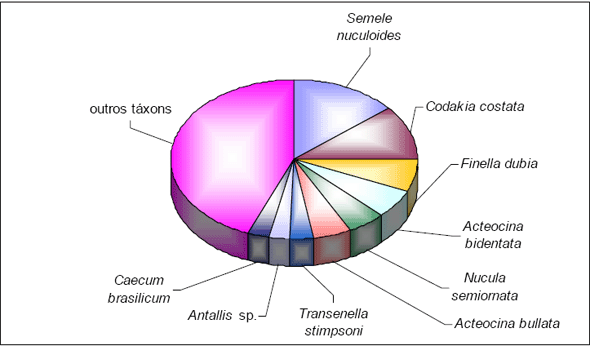
A abundância relativa dessas espécies variou entre as estações de coleta; assim, a espécie Nucula semiornata dominou na estação 1, enquanto Semele nuculoides foi a mais abundante nas estações 2 a 4 (Tabela 111). Codakia costata foi dominante nas estações 5 a 10.
Tabela 111 - Abundância relativa (%) das principais espécies nas estações de coleta.
Clique aqui para visualizar a tabela 111
A análise de variância da densidade indicou existir diferenças significativas entre as estações de coleta e na variação de densidade, entre as estações, pôde ser observado que a Piraquara de Fora apresenta um núcleo central de baixa densidade (estações 4 a 8), cujos valores vão aumentando à medida que se sai desta área (estações 3, 9 e 10).
Figura 128 - Densidade média da fauna total nas estações de coleta.
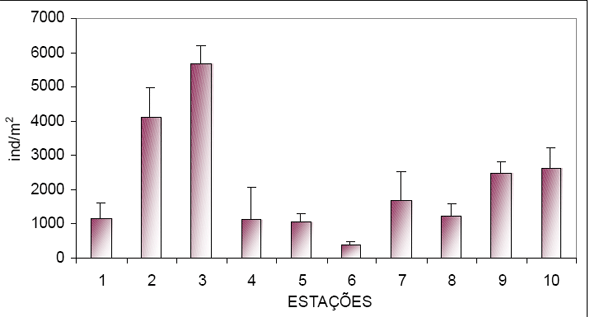
Deve ser ressaltado que entre as estações que formam o núcleo central, a estação 7 (Z4), localizada imediatamente à frente do descarte, foi a que apresentou a densidade mais alta podendo, este fator, estar ligado ao pequeno deslocamento na posição inicial da estação. A baixa densidade da estação 1 provavelmente está relacionada a um hidrodinamismo mais alto, que se refletiu no melhor selecionamento dos grãos do sedimento.
Número de táxons e diversidade
De modo geral, o padrão de distribuição desses dois índices foi muito semelhante e se aproximou bastante do obtido para a densidade (Figura 129 e Figura 130). O núcleo de baixos valores foi basicamente o mesmo, mas a maior diferença foi a incorporação da estação 3 neste núcleo.
Figura 129 - Numero de táxons presentes nas estações de coleta.
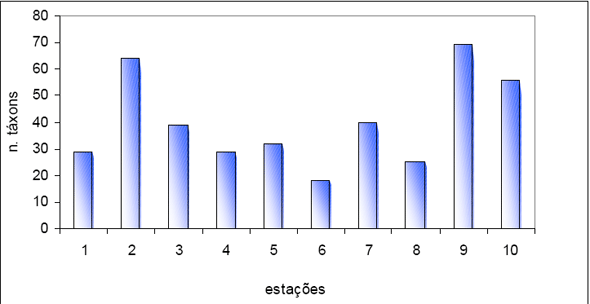
Figura 130 - Diversidade encontrada nas estações de coleta.
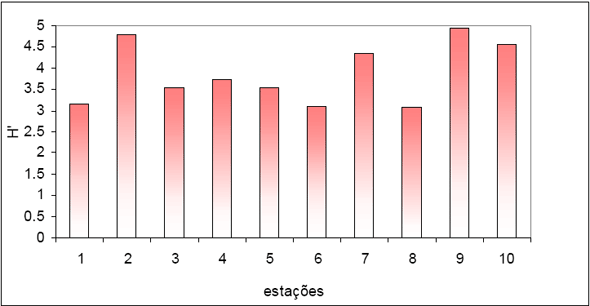
Equidade e dominância
A equitabilidade apresentou valores semelhantes para todas as estações, indicando que a densidade de organismos encontrada nas diversas estações está distribuída de forma regular pelos diversos táxons, o que também pode ser observado pela baixa dominância encontrada. Em locais extremamente estressados ocorre, geralmente, o predomínio de táxons oportunistas (k-estrategistas), trazendo desequilíbrio à estrutura da comunidade (Clarke & Warwick, 1994).
Baseando-se nos índices biológicos, podemos definir um núcleo central representado pelas estações 4 a 8 no qual se encontram os menores valores dos índices biológicos (Figura 131). É importante destacar que dentro desse núcleo central, a estação 7 (Z4) apresentou valores superiores às demais (4, 5, 6 e 8), o que poderia ser explicado, como já comentado anteriormente, pelo pequeno deslocamento na posição inicial desta estação. Apesar de pequeno, o deslocamento pode ter levado a uma amostragem de uma área limítrofe desse núcleo central, o que justificaria, inclusive, a diferença encontrada na classificação granulométrica dessa estação (areia muito fina) em relação às outras do núcleo central (silte).
Figura 131 - Representação, baseada nos índices biológicos, do núcleo central e da região adjacente de estações.
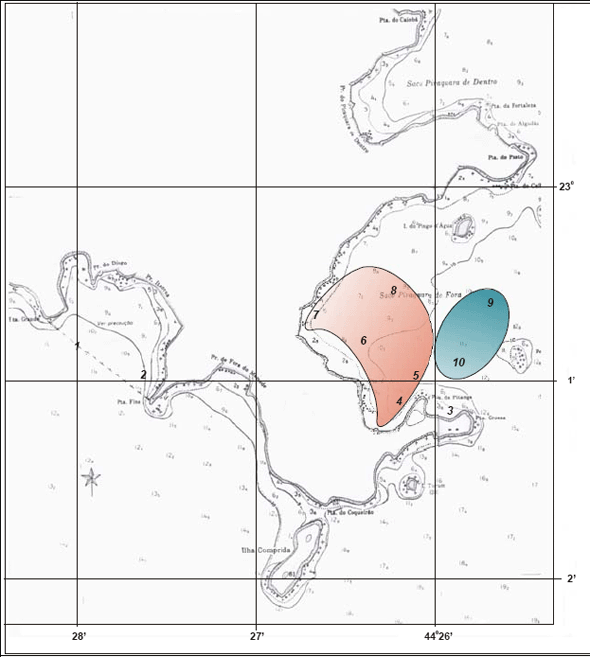
Núcleo adjacente ao central foi formado pelas estações 9 e 10, que se caracteriza pelos maiores valores dos índices biológicos. Associada a este núcleo encontra-se a estação 2 que mesmo localizada em Itaorna, apresenta características semelhantes com às da estação 3 como, por exemplo, a composição sedimentológica. Já a estação 1, também localizada em Itaorna, apresentou-se isolada das demais, o que pode estar associado ao intenso hidrodinamismo local.
Similaridade
A análise de grupamento evidenciou a formação de 3 grupos de estações e uma amostra isolada, no caso a estação 1 (Figura 132). O primeiro grupo (I) é formado pelas estações 9 e 10, com a estação 7 um pouco afastada. O segundo grupo (II) é formado pelas estações 2 e 3 e o terceiro grupo (III) pelas estações 4, 5, 6 e 8. A análise de significância das similaridades (ANOSIM) indicou que a formação destes grupos de estações ocorre de forma significativa (R = 0,94; p < 0.1).
Figura 132 - Dendrograma resultante da aplicação do índice de Bray-Curtis aos valores obtidos para a densidade dos táxons encontrados nas estações de coleta.
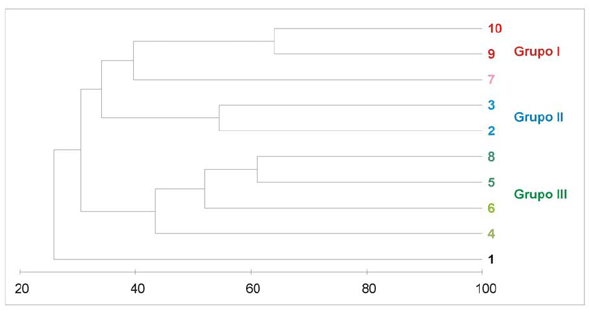
Na Tabela 112 estão relacionadas as espécies que são, significativamente, representativas dos grupos de estações observados a partir da análise de similaridade.
Tabela 112 - Valores significativos do índice de espécies indicadoras de Dufrêne & Legendre (1997) para as espécies encontradas neste estudo.
Clique aqui para visualizar a tabela 112
O padrão da estrutura de comunidade observado na Figura 132 é semelhante ao obtido na análise de grupamento realizada com os parâmetros granulométricos. Esta relação mostrou-se significativa pela análise de correlação de Spearman entre a matriz de similaridade da fauna e a matriz de distância dos parâmetros granulométricos (rs = 0.64; p < 0,1).
Observando o padrão de estrutura da comunidade por meio de análise de ordenação não métrica multimensional (MDS), pôde-se verificar a sua associação às características granulométricas como, por exemplo, o tamanho médio dos grãos (Figura 133).
Figura 133 - Análise multidimensional (MDS) baseada na matriz de similaridade obtida a partir da densidade dos táxons encontrados nas estações de coleta.
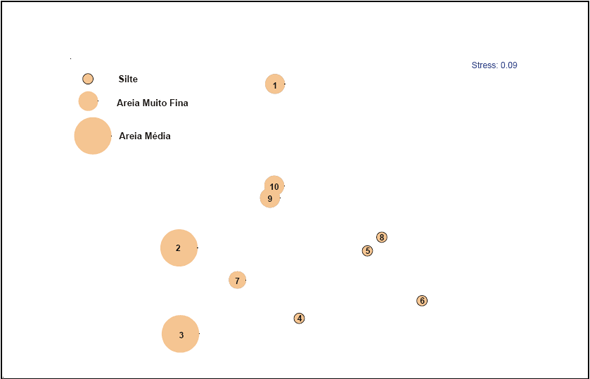
Avaliação Temporal
Nesta avaliação foram utilizados os dados de abundância dos grandes grupos e das espécies indicadas para o monitoramento iniciado em 1982 e obtidos durante os meses de abril e maio. Estes dados estão disponibilizados nos diversos relatórios anteriores. Para se evitar problemas metodológicos como, por exemplo, o uso de diferentes amostradores, os dados aqui apresentados se referem à abundância relativa dos diferentes grupos e espécies indicadas. As áreas de amostragem utilizadas foram: estação fixa Z3 (estação 2 localizada em Itaorna); estação fixa Z4 (estação 7 localizada no Saco Piraquara de Fora, em frente ao efluente) e quadrante I, representado por um conjunto de estações localizadas no Saco Piraquara de Fora (estações 4, 5, 6 e 10), também denominada área B de Impacto.
Nas estações acima indicadas foram obtidos 58 táxons nos meses de abril e maio de 1980 e 144 táxons em 2002, sendo 47 táxons comuns aos dois estudos.
Analisando a variação temporal dos grandes grupos, percebe-se que o grupo Mollusca foi o dominante em todas as áreas desde do início dos estudos (Figura 134, Figura 135 e Figura 136). No entanto, a abundância desse grupo no quadrante I (área de impacto) aumentou ao longo do período avaliado, enquanto que o grupo Echinodermata sofreu uma redução nos seus valores de abundância ao longo do tempo nesta mesma área. Este padrão não foi observado para as estações Z3 e Z4, onde, de forma geral, os valores de abundância de Mollusca se mantiveram como anteriormente.
Figura 134 - Abundância relativa dos grandes grupos, no quadrante I, no período de 1980 a 2002.
Figura 135 - Abundância relativa dos grandes grupos, na estação Z4, no período de 1983 a 2002.
Figura 136 - Abundância relativa dos grandes grupos, na estação Z3, no período de 1983 a 2002.
As espécies indicadas para o monitoramento pela UFRJ (1981) encontram-se relacionadas na Tabela 113. A partir dessa indicação, foram iniciados, em 1982/1983 (Furnas, 1983), estudos de acompanhamento das espécies, estudos, esses, mantidos até 2001 (ELETROBRÁS TERMONUCLEAR , 2002).
Tabela 113 - Espécies indicadas para o monitoramento pela UFRJ (1981).
Clique aqui para visualizar a tabela 113
Inicialmente, as espécies de Polychaeta não foram avaliadas nesse acompanhamento devido a problemas taxonômicos que seriam posteriormente solucionados (Furnas, 1983) mas, nas avaliações posteriores, não há nenhuma referência a soluções taxonômicas nem, mesmo, uma justificativa para a não inclusão dessas espécies no programa de monitoramento. Nesta avaliação, a espécie Laonice cirrata esteve presente com pequenos valores de abundância, não sendo considerada uma espécie importante quanto a esta variável. A espécie Sthenolepis oculata não foi registrada na campanha realizada este ano.
Outra espécie, Leptochela serratorbita (Crustacea), também não foi acompanhada após 1983, apesar de não existir qualquer comentário sobre a retirada desta espécie do monitoramento em qualquer dos relatórios técnicos disponibilizados. Supõe-se que o fato pode estar relacionado à redução da freqüência e/ou densidade dessa espécie do ano de 1980 para 1982/1983. Na avaliação deste ano (2002), esta espécie também não foi registrada em qualquer das estações de coleta.
Considerando as espécies realmente utilizadas no monitoramento, podemos observar que Codakia costata, Nucula semiornata, Tellina sandix e Edwardsia sp. continuam sendo importantes em termos de abundância. No entanto, as espécies Corbula caribaea, Chione paphia, Dentalium gouldii, Pitar sp.e Dentalium sp. não estiveram presentes nas estações de coleta localizadas no quadrante I (estações 4, 5, 6 e 10) nem nos pontos fixos Z3 (estação 2) e Z4 (estação 7).
Quanto ao padrão temporal na abundância das espécies indicadas, verifica-se que a espécie Codakia costata apresentou um aumento nos seus valores de abundância no quadrante I; já as demais espécies como, por exemplo, Nucula semiornata e Tellina sandix,mantiveram os seus valores ao longo do tempo (Figura 137).
Figura 137 - Abundância relativa das espécies indicadas, no quadrante I, no período de 1980 a 2002.
Na estação fixa Z4 (estação 7), apesar das variações ao longo do tempo, Codakia costata apresentou valores neste estudo semelhantes aos do início dos estudos em 1980 (Figura 138).
Figura 138 - Abundância relativa das espécies indicadas, na estação Z4, no período de 1983 a 2002.
A estação Z3 foi a que apresentou as maiores variações nas abundâncias das espécies, especialmente, de Codakia costata, Nucula semiornata e Tellina sandix (Figura 139).
Figura 139 - Abundância relativa das espécies indicadas, na estação Z3, no período de 1983 a 2002.
Considerações Finais sobre Zoobentos
A sedimentologia da área de estudo apresentou-se, predominantemente, fina e muito mal selecionada a moderadamente selecionada, sendo este resultado, de modo geral, muito semelhante aos estudos realizados em 1980/81 e 2002.
As estações apresentaram, entre si, diferenças nas características granulométricas.
Foi encontrada uma densidade média de 2151 org/m2 na área estudada e 144 táxons que representaram 12 grandes grupos zoológicos, entre os quais o filo Mollusca foi o mais abundante em todas as estações de coleta.
Nove espécies de Mollusca foram as mais abundantes e responsáveis por 60% da fauna total: Semele nuculoides, Codakia costata, Finella dubia, Acteocina bidentata, Nucula semiornata, Acteocina bullata, Transenella stimpsoni, Antallis sp.e Caecum brasilicum.
O segundo grupo, em importância, foi Annelida e as espécies mais abundantes desse grupo foram: Polygordius sp., Hemipodus sp., Prionospio sp.1, Magelona sp., Haploscoloplos sp. e Magelona cincta.
O terceiro grupo mais abundante foi Crustácea, com as espécies de anfípodos Ampelisciphotis podophtalma, Tiborunella viscana e Ampelisca cristata sendo as mais importantes desse grupo.
O quarto grupo foi Cnidaria com uma única espécie importante Edwardsia sp.
A partir dos índices biológicos foi possível definir um núcleo central, na Piraquara de Fora, representado pelas estações 4 a 8, no qual se encontram os menores valores desses índices. Apesar de se localizar nesse núcleo central, a estação 7 apresentou valores superiores às demais, podendo, essa diferença encontrada, estar associada à composição granulométrica desta estação.
O núcleo adjacente ao central foi formado pelas estações 9 e 10, que se caracteriza pelos maiores valores dos índices biológicos. Associado a este núcleo encontra-se a estação 2 que, mesmo localizada em Itaorna, apresentou características semelhantes à estação 3. A estação 1, também localizada em Itaorna, apresentou isolada das demais, o que pode estar associado ao hidrodinamismo intenso dessa estação.
Na avaliação temporal foram obtidos 58 táxons nos meses de abril e maio de 1980 e 144 táxons em 2002, sendo 47 táxons comuns aos dois estudos.
A análise da variação temporal dos grandes grupos mostrou que o grupo Mollusca foi o dominante, em todas as áreas, desde do início dos estudos. Mas, especialmente, na área denominada quadrante I, aumentou sua abundância ao longo do período avaliado, enquanto que o grupo Echinodermata sofreu uma redução nos seus valores de abundância, ao longo do tempo, nessa mesma área.
Quanto às espécies utilizadas no monitoramento, observou-se que Codakia costata, Nucula semiornata, Tellina sandix e Edwardsia sp. continuam sendo importantes em termos de abundância. No entanto, as espécies Corbula caribaea, Chione paphia, Dentalium gouldii, Pitar sp.e Dentalium sp. não estiveram presentes nas estações de coleta localizadas no quadrante I, nem nos pontos fixos Z3 e Z4.
A análise temporal da abundância das espécies indicadas mostrou que Codakia costata apresentou um aumento nos seus valores de abundância, no quadrante I; já as demais espécies como, por exemplo, Nucula semiornata e Tellina sandix mantiveram os seus valores ao longo do tempo. Apesar das variações ao longo do tempo, Codakia costata, na estação fixa Z4,apresentou valores neste estudo semelhantes aos encontrados em 1980/81.
7.5. Macroinvertebrados Terrestres e Aquáticos - topo
O levantamento de dados existentes, elaborado a partir de pesquisa bibliográfica, pertinentes aos principais grupos e à área de estudos, considerou, principalmente, estudos faunísticos e listas de espécies publicadas, além de teses e dissertações referentes à região litorânea do sul do Estado do Rio de Janeiro e áreas fronteiriças de mesma fisionomia.
Preliminarmente, um total de 242 táxons estão registrados para a área estudada, distribuídos da seguinte forma: Decapoda (9 spp.), Ephemeroptera (5 spp.), Odonata (36 spp.), Hemiptera (19 spp.), Homoptera (11 spp.), Coleoptera (45 spp.) Hymenoptera (5 spp.), Mecoptera (1 sp.), Trichoptera (26 spp.), Lepidoptera (15 spp.) e Diptera (70 spp.).
Crustacea - Decapoda
Dentre os Decapoda, apenas as famílias Atyidae, Palaemonidae e Trichodactylidae estão representadas na rede fluvial no Estado do Rio de Janeiro (Rodriguez, 1981, Coelho et al. 1985). Os Atyidae e Palaemonidae são camarões bentônicos. A família Atyidae, quase completamente confinada às águas doces e aos habitats subterrâneos, está representada pelos gêneros Atya, de ocorrência pantropical e Potimirim, endêmico na região neotropical. Nos Papaemonidae, de ampla distribuição no mundo, incluem-se os camarões de rio do gênero pantropical Macrobrachium, assim como outros gêneros de água doce e marinhos. Trichodactylidae é uma família de caranguejo estritamente de água doce, habita os grandes rios e lagos das terras baixas das grandes bacias continentais da América do Sul a leste dos Andes (Rodriguez,1992).
O conhecimento da fauna de decápodos dulciaqüícola do Rio de Janeiro, é ainda muito fragmentado, recebendo atualmente atenção mínima dos pesquisadores. Villalobos (1959) descreveu uma nova espécie para o Brasil, Potimirim brasiliana, proveniente do rio Ariró (Angra dos Reis). Os camarões das famílias Palaemonidae e Atyidae foram objeto de estudo para a dissertação de mestrado de Fonseca (1995). Esta representa a mais atualizada contribuição das espécies de camarões de água doce, para o Estado.
Tabela 114 - Espécies de crustáceos decápodos registrados para a bacia hidrográfica: contribuinte à baía de Ilha Grande.
| Família |
Espécie |
Nome Vulgar |
Altitude (m) |
Atyidae |
Atya scabra |
Camarão da pedra |
0 a 400 |
| |
Potimirim glabra |
--- |
0 a 100 |
| |
P. potimirim |
--- |
0 a 100 |
Palaemonidae |
Palaemon pandaliformes |
Camarão potitinga |
0 a 100 |
| |
Macrobrachium acanthurus |
Camarão canela |
0 a 250 |
| |
M. carcinus |
Camarão pitu |
0 a 200 |
| |
M. olfersii |
Camarão aratanha |
0 a 100 |
| |
M. potiuna |
Camarão sossego |
Até 120 |
| |
|
Camarão preto |
|
Trichodactylidae |
Trichodactylus fluviatilis |
Pitangola |
0 a 1.000 |
Insecta - Ephemeroptera
Cerca de trinta espécies nominais de Ephemeroptera estão registradas para o Estado do Rio de Janeiro, a maioria das quais com ocorrência conhecida na Região Serrana. Para o Litoral Sul, estão registradas seis espécies. Levando-se em conta que o Litoral Sul apresenta trechos ainda bem preservados de Mata Atlântica, ricos em um dos habitats preferenciais de ninfas de Ephemeroptera, os ambientes lóticos ritrais de pequeno a médio porte, é de se esperar que, com novas coletas e estudos sistemáticos, tal número seja em muito acrescido. Novas coletas provavelmente revelarão a ocorrência na região das famílias Baetidae, Leptohyphidae e Oligoneuriidae, bem representadas em corpos ritrais fluminenses. As referências para a área de estudo são Da Silva (2002) e Pereira & Da Silva (1990).
Tabela 115 - Lista dos Ephemeroptera com ocorrência registrada o litoral sul do Estado do Rio de Janeiro.
| Família |
Espécie |
| Euthyplociidae |
Campylocia bocainensis Pereira & Da-Silva, 1990 |
| Leptophlebiidae |
Askola froehlichi Peters, 1969 |
| |
Farrodes carioca Domínguez, Molineri & Peters, 1996 |
| |
Massartella brieni (Lestage, 1924) |
| |
Miroculis froehlichi Savage & Peters, 1983 |
| |
Thraulodes itatiajanus Traver & Edmunds Jr, 1967 |
Insecta - Odonata
Existem poucos trabalhos de levantamento da fauna de insetos ocorrentes no Brasil, e o Estado do Rio de Janeiro não se trata de uma exceção. No caso dos Odonata existem listagens apenas para o Parque Nacional de Itatiaia (Santos, 1970), para a Restinga do Recreio dos Bandeirantes (Santos, 1965), para a Restinga de Maricá (Carvalho, 1991), e para algumas ilhas costeiras do Estado (Carvalho & Pujol-Luz, 1992). É possível que hajam trabalhos destinados ao tratamento de algumas espécies isoladas em que material coletado no município de Angra dos Reis tenha sido citado, mas tais informações são muito esparsas e difíceis de serem resgatadas. Carvalho & Pujol-Luz (1992) registraram 36 espécies de Odonata para a Ilha Grande, Angra dos Reis, RJ, que estão listadas abaixo.
Tabela 116 - Lista dos Odonata registrados para Ilha Grande, Angra dos Reis, segundo Carvalho & Pujol-Luz (1992).
Clique aqui para visualizar a tabela 116
Praticamente todas essas espécies apresentam ampla distribuição no Sudeste do Brasil, sendo características de áreas de Mata Atlântica. A espécie Mecistogaster asticta, ocupante de zonas umbrófilas, citadas como rara nesse mesmo artigo, está também registrada como espécie ameaçada de extinção (Bergallo et al., 2000).
Carvalho & Nessimian (1998) listam 77 gêneros distribuídos em doze famílias ocorrentes no Estado do Rio de Janeiro, comentando a existência de pelo menos 245 espécies nesse estado. Material imaturo de cinco espécies ocorrentes em Angra dos Reis está registrado, sendo que essas estão citadas no trabalho de Carvalho & Pujol-Luz (1992).
Insecta - Hemíptera
Pouca informação acerca dos Hemiptera terrestres está disponível para a área de estudos. Contudo, mais pesquisas são necessárias, dada a importância da ordem, sua diversidade e a grande quantidade de artigos isolados e material depositado. No que concerne ao conhecimento da diversidade dos Hemiptera aquáticos no Estado do Rio de Janeiro, pouco estudos foram realizados até o momento. Assim, os registros faunísticos desse estado, bem como os estudos ecológicos sobre os diversos habitats desse grupo são escassos, com somente quatro registros para a área de estudo.
Tabela 117 - Lista dos Hemiptera registrados no litoral sul do Estado do Rio de Janeiro, especialmente na região de Angra dos Reis.
| Família |
Espécie |
| Aradidae |
Asterocoris australis Drake e Harris |
| Miridae |
Crassicornus pulchrus Carvalho, 1945 |
| |
Derophthalma fluminensis Carvalho, 1944 |
| |
Euchilocoris hahni (Stal, 1860) |
| |
Guanabarea angrensis Carvalho 1948 |
| |
Hyaliodes wygodzinskyi Carvalho, 1945 |
| |
Parachius rufovittatus Carvalho, 1944 |
| |
Paramixia carmelitana (Carvalho, 1948) |
| |
Phytocoris citrinoides Carvalho & Fontes, 1970 |
| |
P. subvittatus (Stal, 1860) |
| |
Polymerus testaceipes (Stal, 1860) |
| |
Halticus bractatus (Say, 1832) |
| |
Neela lutescens (Stal, 1860) |
| |
Prepos zetterstedti var. pyrrhomelaemus (Stal, 1860) |
| |
Pycnoderes leucopus (Stal, 1860 ) |
| Naucoridae |
Limnocoris brasiliensis |
| |
L. nigropunctatus Montandon, 1909 |
| Nepidae |
Ranatra costalimai |
| Ochteridae |
Ochterus perbosci |
Insecta - Homoptera
Dentre os Homoptera, na lista preliminar dos cicadelídeos do Brasil (Zanol & de Menezes, 1982), ocorrem na região de Angra dos Reis-RJ onze espécies de seis subfamílias. Dentre essas espécies, oito são conhecidas somente dessa região (localidade-tipo), podendo ser consideradas endêmicas. Referências para grupos como Cicadidae, Membracidae e Cercopidae não foram encontradas, mas certamente o número de espécies ocorrentes em Angra dos Reis é muito maior.
Tabela 118 - Lista dos cicadelídeos ocorrentes em Angra dos Reis-RJ, segundo Zanol & de Menezes (1982). Espécies endêmicas dessa região estão assinaladas por um asterisco (*).
| Subfamília |
Espécie |
Agalliinae |
Agallia cobera Kramer, 1964* |
| |
A. cucata Kramer, 1964* |
| |
Agalliopsis zenestra Kramer, 1964 |
Cicadellinae |
Platygonia angrana Young, 1977* |
Deltocephalinae |
Sanctanus lepidellus (Stål, 1862) |
| |
S. vulpinus Kramer, 1963* |
Gyponinae |
Hecalapona (Carapona) ferosa DeLong & Freytag, 1975* |
| |
Polana (Parvulana) bidens DeLong & Freytag, 1972* |
Iassinae |
Pachyopsis similis Kramer, 1963* |
Nirvaninae |
Krocobella colotes Kramer, 1964* |
| |
Neonirvana hyalina Oman, 1936 |
Insecta - Coleoptera
Embora seja o maior grupo animal em número de espécies, há poucos registros para a área do empreendimento. Os dados obtidos foram retirados de listas e coleções. Contudo, as espécies relacionadas abaixo não refletem absolutamente o potencial da região e sim a quase ausência de coletas específicas para o grupo.
Tabela 119 - Espécies e gêneros de Coleoptera com ocorrência registrada para o litoral sul do Estado do Rio de Janeiro.
Clique aqui para visualizar a tabela 119
Insecta - Mecoptera
Somente uma espécie está registrada para a área, por Zikán & Vygodzinsky (1948) (Tabela 120).
Insecta - Hymenoptera
Quatro espécies estão registradas para a área, por Zikán & Vygodzinsky (1948) (Tabela 120).
Tabela 120 - Tipos de Insecta das ordens Mecoptera e Hymenoptera coligidos em Angra dos Reis (Zikán & Wygodzinsky, 1948)
Ordem |
Família |
Espécie |
Hymenoptera |
Cynipidae |
Epicoela angrensis Borgmeier |
| |
|
Tropideucoila angrensis Borgmeier |
| |
Formicidae |
Basiceros squamifer Borgmeier |
| |
|
Eciton (Neivamyrmex) porrectognathum Borgmeier |
Mecoptera |
Bittacidae |
Bittacus angrensis Lopes & Mangabeira |
Insecta - Trichoptera
Com relação à ordem Trichoptera, no Estado Rio de Janeiro foram registradas 50 espécies segundo o Catalogo de Flint et al. (1999) e bibliografia especializada. No Rio de Janeiro, o Município de Nova Friburgo apresenta o maior número de citações, e além dele estão: Teresópolis, Cachoeiras de Macacú, Rio Claro, Parque Nacional Tijuca e Parque Nacional Itatiaia.
A região de Angra dos Reis apresenta quatro citações. No estado também foram realizados estudos na taxonomia e biologia das larvas: Guahyba (1981) No Parque Nacional Tijuca e Huamantinco & Nessimian (1999, 2000) no município de Teresópolis. Carrijo et al. (2001) listam alguns gêneros presentes no litoral sul do Estado do Rio de Janeiro.
Tabela 121 - Espécies e gêneros de Trichoptera com ocorrência registrada para o litoral sul do Estado do Rio de Janeiro.
Família |
Gênero/Espécie |
Calamoceratidae |
Phylloicus sp. |
Glossosomatidae |
Mortoniella sp. |
Helicopsychidae |
Helicopsyche sp. |
Hydrobiosidae |
Atopsyche sp. |
Hydropsychidae |
Blepharopus sp. |
| |
Leptonema sp |
| |
Leptonema pallidum |
| |
Macronema sp. |
| |
Macronema fulvum |
| |
Macrostemum sp. |
| |
Smicridea sp. |
Hydroptilidae |
Ochrotrichia sp. |
| |
Scactobiella sp. |
Leptoceridae |
Atanatolica sp. |
Leptoceridae |
Grumichella sp. |
| |
Nectopsyche sp. |
| |
Notalina sp. |
| |
Oecetis sp. |
| |
Triplectides sp. |
Odontoceridae |
Barypenthus sp. |
| |
Mariliasp. |
Philopotamidae |
Chimarra beckeri |
| |
Chimarra camura |
| |
Wormaldia sp. |
Polycentropodidae |
Polycentropus sp. |
Sericostomatidae |
Grumicha sp. |
Insecta - Lepidoptera
Na porção da Mata Atlântica, situada nos arredores do Município de Angra dos Reis, temos um elevado número de espécies de insetos da Ordem Lepidoptera. Esta diversidade é decorrente do fato de ser o município vizinho do Parque Nacional da Serra da Bocaina, e também, por ele apresentar diversas áreas de conservação.
Tabela 122 - Lista dos Lepidoptera endêmicos de Mata Atlântica, ocorrentes na região de Angra dos Reis.
Família |
Espécie |
Nymphalidae |
Agrias claudina claudina (Godart, 1824). |
| |
Dasyothatalma rusina (Godart, 1824) |
| |
Heliconius besckei Ménétries, 1857 |
Papilionidae |
Battus polystictus galenus (Fruhstorfer, 1907) |
| |
Heraclides himeros (Hopffer, 1856) |
| |
Mimoides lysithous lysithious (Hubner, 1821) |
| |
Parides ascanius, (Cramer, 1776) |
| |
Parides tros (Fabricius, 1793) |
| |
Parides zacynthus zacynthu (Fabricius, 1793) |
| |
Protesilaus molops megalurus (Rothschild & Jordan, 1906) |
| |
Protrographium asius (Fabricius, 1782) |
Insecta - Diptera
Com relação à fauna de Diptera do litoral sul fluminense e proximidades, os principais grupos não estão contemplados. Zikán & Vygodzinsky (1948) listaram algumas espécies de Stratiomyiidae,
Ephydridae e Sarcophagidae para a área do empreendimento; Guimarães et al. (2000) estudaram a fauna de Culicidae do Parque Estadual da Serra do Mar, Picinguaba, Ubatuba, SP onde listaram a ocorrência de 55 espécies A família Simuliidae está representada por seis espécies, das quais Simulium (Chirostibia) pertinax e S.
(Psaroniocompsa) incrustatum são os mais abundantes da região. Contudo, o número de espécies desta família é baixo com relação ao existente na Região Neotropical (330) (Crosskey, & Howard, 1997 apud Coscarón & Coscarón-Arias, 2002). Com relação à família Ceratopogonidae, no parte dos estudos desta família no Brasil foram feitos na Fundação Oswaldo Cruz com material depositado na Coleção Entomológica desta instituição com oito gêneros e 58 espécies (Felippe-Bauer & Oliveira, 2001), ocorrendo no Estado do Rio de Janeiro 30 espécies distribuídas em seis gêneros:, Downeshelea (5), Monohelea (2); Culicoides (15); Palpomyia (1); Focipomyia (5); Dasyhelea (2). Destas, somente duas espécies estão registradas para o litoral sul do estado.
Outro grupo de grande diversidade, Chironomidae, com cerca de 100 gêneros presentes no Estado do Rio de Janeiro (Nessimian et al. 2001), ainda não possui qualquer registro para a área de estudos.
Tabela 123 - Espécies e gêneros de Diptera com ocorrência registrada para o litoral sul do Estado do Rio de Janeiro.
| Família |
Espécie |
Ceratopogonidae |
Culicoides tavaresi Fellipe-Bauer & Wirth, 1988 |
| |
Dasyhelea pulsilla Lutz, 1913 |
Ephydridae |
Stenochthera nigricoxa Cresson |
Sarcophagidae |
Sarcophaga crispina Lopes |
| |
S. expuncta Lopes |
Simuliidae |
Simulium (Psaroniocompsa) angrense |
| |
Simulium (Inaequalium) clavibranchium |
| |
Simulium (Psaroniocompsa) incrustatum |
| |
Simulium (Chirostibia) pertinax |
| |
Simulium (Inaequalium) subnigrum |
| |
Simulium (Inaequalium) travassosi |
Stratiomyiidae |
Berisargus fasciatipennis Lindner |
| |
B. Lopesi Lindner |
| |
Glariopsis brasiliana Lindner |
| |
Neochorisops brasiliensis Lindner |
Tabela 124 - Espécies de Culicidae (Diptera) encontrados por Guimarães et al. (2000) na área do Parque Estadual da Serra do Mar , Picinguaba, Ubatuba, SP.
Clique aqui para visualizar a tabela 124
O levantamento da entomofauna terrestre e dos macroinvertebrados aquáticos foi realizado em três localiadades escolhidas na área de influência do empreendimento, doravante denominadas Bracuí, entre 44o 23’ e 44o 25’ W e 22o 54’ e 23o 57’ S; Praia Brava, entre 44o 26’ e 44o 29’ W e 23o 58’ e 23o 01’ S e Mambucaba, entre 44o 30’ e 44o 35’ W e 22o 58’ e 23o03’ S, nas quais foram realizadas coletas em treze pontos.
Além destas, para a fauna aquática, foram utilizados dados referentes a dois pontos, considerados controle, o rio Bonitinho, um tributário do rio Bracuí na localidade Bracuí e o rio Caputera, em Jacuecanga. Os pontos de amostragem estão assinalados na Tabela 125, com seus respectivos números e códigos utilizados nas tabelas e na Figura 140.
Tabela 125 - Pontos de amostragem na área de influência da Usina Angra 3, com respectivos números e códigos utilizados.
| No |
Localidade |
Ponto de Amostragem |
Código |
| 01 |
Mambucaba |
Trecho potamal alto do rio Mambucaba |
MA |
| 02 |
Mambucaba |
Trecho ritral baixo do rio da Cachoeirinha |
CCH |
| 03 |
Mambucaba |
Trecho ritral baixo do rio Água branca |
ABR |
| 04 |
Mambucaba |
Poças marginais ao rio Água Branca |
POÇ |
| 05 |
Mambucaba |
Trecho ritral baixo do rio Guariri |
GUA |
| 06 |
Mambucaba |
Trecho ritral baixo do rio Itapicu (Perequê) |
|
| 07 |
Praia Brava |
Trecho ritral baixo de tributário do rio Cachoeira Brava |
PB1 |
| 08 |
Praia Brava |
Trecho ritral baixo do rio Cachoeira Brava |
PB2 |
| 09 |
Praia Brava |
Trecho ritral baixo de córregos próximos à Usina |
USI |
| 10 |
Bracuí |
Açude marginal à rodovia BR101 próximo ao rio Grataú |
AÇU |
| 11 |
Bracuí |
Condomínio Porto Bracuí, na seção final do rio Bracuí |
|
| 12 |
Bracuí |
Trecho potamal do rio Bracuí |
BRP |
| 13 |
Bracuí |
Trecho ritral baixo do rio Bracuí |
BRR |
| 14 |
Bracuí |
Trecho ritral baixo de rio Bonitinho, tributário do rio Bracuí |
BTR |
| 15 |
Jacuecanga |
Trecho ritral baixo do rio Caputera |
CAP |
Figura 140 - Localização dos pontos de coleta na área de influência da Usina Angra 3 (Fonte: CD Brasil - Embrapa).
Os pontos de coleta foram avaliados quanto ao grau de preservação da cobertura vegetal original e uso da terra, sendo atribuídos valores relativos de 1 (muito perturbada) a 5 (sem perturbação). Da mesma forma os rios amostrados foram caracterizados fisionomicamente (cobertura vegetal, uso da terra, substrato dominante). Para a caracterização do estado ambiental dos rios foi utilizada uma versão modificada do inquérito ambiental de Petersen (1992). (Tabela 126 e Tabela 127).
Tabela 126 - Cobertura vegetal e uso da terra nos pontos de amostragem na área de influência da Usina Angra 3.(OBS.: 1 - muito perturbada; 5 - sem perturbação).
| No |
Localidade |
Cobertura vegetal e uso da terra |
Descrição |
Pontuação |
| 01 |
Mambucaba |
Floresta, capoeira, culturas permanentes, habitações isoladas |
3 |
| 02 |
Mambucaba |
Floresta, capoeira, culturas permanentes, aglomerações habitacionais |
2 |
| 03 |
Mambucaba |
Floresta, capoeira, culturas permanentes, habitações isoladas |
3 |
| 04 |
Mambucaba |
Floresta, capoeira, culturas permanentes, habitações isoladas |
3 |
| 05 |
Mambucaba |
Floresta, capoeira, culturas permanentes, sítios, habitações isoladas |
3 |
| 06 |
Mambucaba |
Floresta, capoeira, pastagem, sítios, área periurbana |
2 |
| 07 |
Praia Brava |
Floresta, capoeira, proximidade com rodovia pavimentada |
3 |
| 08 |
Praia Brava |
Floresta, capoeira, proximidade com rodovia pavimentada |
3 |
| 09 |
Praia Brava |
Floresta, capoeira, construções civis, linha de transmissão, proximidade com rodovia pavimentada |
2 |
| 10 |
Bracuí |
Capoeiras e áreas urbanizadas |
1 |
| 11 |
Bracuí |
Pastagem, capoeira, sítios e fazendas proximidade com rodovia pavimentada |
1 |
| 12 |
Bracuí |
Pastagem, capoeira, sítios, aglomerações habitacionais |
1 |
| 13 |
Bracuí |
Floresta, capoeira, sítios, habitações isoladas |
3 |
| 14 |
Bracuí |
Floresta, capoeira, sítios, habitações isoladas |
4 |
| 15 |
Jacuecanga |
Floresta, capoeira, sítios, habitações isoladas |
4 |
Tabela 127 - Avaliação fisionômica dos trechos de rios amostrados na área de influência de Angra 3, segundo o protocolo modificado de Petersen (1992).
| No |
Pontos de amostragem |
Código |
Petersen |
| Pontução |
Avaliação |
| 01 |
Trecho potamal alto do rio Mambucaba |
MA |
271 |
B |
| 02 |
Trecho ritral baixo do rio da Cachoeirinha |
CCH |
356 |
MB |
| 03 |
Trecho ritral baixo do rio Água branca |
ABR |
291 |
B |
| 05 |
Trecho ritral baixo do rio Guariri |
GUA |
291 |
B |
| 07 |
Trecho ritral baixo de tributário do rio Cachoeira Brava |
PB1 |
365 |
MB |
| 08 |
Trecho ritral baixo do rio Cachoeira Brava |
PB2 |
385 |
E |
| 09 |
Trecho ritral baixo de córregos próximos à Usina |
USI |
151 |
P |
| 12 |
Trecho potamal do rio Bracuí |
BRP |
133 |
P |
| 13 |
Trecho ritral baixo do rio Bracuí |
BRR |
380 |
E |
| 14 |
Trecho ritral baixo de rio Bonitinho, trib. do rio Bracuí |
BTR |
390 |
E |
| 15 |
Trecho ritral baixo do rio Caputera |
CAP |
420 |
E |
B-bom; E - excelente, MB - muito bom; P - pobre.
Em todos os pontos de coleta, exceto os pontos 14 e 15, no período diurno (durante cerca de 10 horas), os insetos aéreos foram coletados em pleno vôo e na vegetação.
Para a captura de insetos em vôo foram utilizadas redes entomológicas e para os que se encontravam na vegetação foram utilizados sugadores entomológicos e câmaras mortíferas, contendo éter etílico como substância anestésica. Os insetos ocorrentes no solo e em troncos caídos foram capturados diretamente através de catação manual. Foram empregadas ainda armadilhas de interceptação do tipo Malaise, montadas e deixadas por um período de 24 horas (pontos 01, 07, 09, 12 e 13). Foram montadas armadilhas para moscas, utilizando-se carne de peixe como isca (pontos 01, 06, 07, 08, 09, 11, 12 e 13). Estas foram deixadas suspensas a 2 m do solo, também, por 24 horas.
Em todas as expedições de coleta foram realizadas amostragens no período noturno. Neste tipo de coleta foi utilizada armadilha do tipo Pensilvânia, com luz branca e ultra violeta. As coletas foram iniciadas no crepúsculo vespertino, meia hora antes do anoitecer, e as armadilhas foram deixadas durante toda a noite, sendo retiradas no dia seguinte. Os pontos de amostragem foram 01, 06, 07, 08, 09, 12 e 13.
Foram realizadas coletas qualitativas de insetos dulçaqüícolas em diversos corpos d’água nas áreas estudas (pontos 01, 02, 03, 04, 05, 07, 08, 09, 10, 12 e 13). A coletas foram realizadas com o emprego peneiras e puças com malha de 1 mm e um amostrador do tipo Surber, com abertura de malha de 200 µm, nos substratos areia, pedra, folhiço retido em área de correnteza e folhiço depositado em área de remanso.
Os insetos aéreos coletados foram, segundo o caso, anestesiados e mortos em éter etílico ou acetato de etila e conservados a seco em camadas de papel ou em envelopes entomológicos, para posterior montagem em laboratório, ou fixados e conservados em álcool etílico a 80%. As libélulas e as borboletas coletadas foram acondicionadas em envelopes entomológicos e deixadas morrer por inanição. Todo o material foi etiquetado, anotando-se a área de proveniência. Os insetos aquáticos foram fixados em álcool etílico a 80%.
No laboratório, os insetos aéreos coligidos foram separados, montados, etiquetados e guardados em caixas entomológicas. O material aquático foi conservado em frascos com álcool etílico a 80% e acondicionado em frascos de vidro, etiquetado com a área de procedência.
A identificação taxonômica dos exemplares coletados foi realizada no nível máximo possível através de literatura específica, comparações com exemplares depositados em coleção e com o auxílio de especialistas. Quando não foi possível determinar a espécie, os exemplares foram separados em morfotipos dentro de um gênero, tribo, subfamília e família.
O material coletado encontra-se depositado nas coleções entomológicas da Universidade Federal do Rio de Janeiro (Instituto de Biologia e Museu Nacional).
O levantamento totalizou nove espécies de Crustacea Decapoda e pelo menos 1200 espécies ou morfótipos de Insecta nas áreas de influência direta e indireta do empreendimento. Desse total, apenas 454 espécies, ou seja, 37,8% foram coletadas nas campanhas.
Figura 141 - Táxons de Insecta e Crustacea Decapoda registrados na região de Angra dos Reis, RJ.
Grande parte dos táxons presentes possui ampla distribuição geográfica e está relacionada à áreas abertas (com alterações na cobertura vegetal original), com ocorrências em todas as ordens amostradas. Algumas espécies encontradas podem ser consideradas periurbanas ou sinantrópicas, como exemplos em Diptera e Lepidoptera. Dentre as ordens de Insecta consideradas, as mais bem representadas nas coletas foram Coleoptera, com 90 espécies; Homoptera, com 89 spp.; e Lepidoptera, com 88 spp. Contudo esses valores, certamente, não retratam a diversidade da região. Esse fato se justifica pela grande falta de conhecimento acerca da entomofauna local, pelo pequeno período de coletas e pela época em que foram realizadas (outono-inverno, período seco). Do material coletado, 272 táxons representam novos registros, não tendo sido, ainda, colecionados ou não constando nas poucas listas ou catálogos que incluam a região de Angra dos Reis. Alguns novos registros para o Estado do Rio de Janeiro foram verificados, como, por exemplo, os gêneros Allenhyphes (Ephemeroptera) e Mexitrichia (Trichoptera). Constatou-se, também, a ocorrência de espécies ainda não conhecidas para a ciência.
A seguir serão apresentados os resultados referentes a cada grupo, com um breve comentário acerca de seu estado de conhecimento.
Crustacea - Decapoda
Os Decapoda estão representados por nove espécies (Tabela 128), na área de influência do empreendimento. Nas campanhas realizadas foram encontrados somente duas de camarões (Atyidae e Palaemonidae) e uma de carangueijo (Trichodactylidae). Segundo Fonseca (1995), a salinidade influencia o padrão de distribuição das espécies de camarão de água doce nas bacias hidrográfica do Rio de Janeiro. As espécies denominadas litorâneas, como Potimirim glabra, requerem uma certa concentração de cloreto de sódio para seu completo desenvolvimento larvar, mantendo-se, assim, próximas à margem do continente. Estas espécies passam as primeiras fases de seu desenvolvimento nos estuários, retornando, posteriormente ao rio. Este fato indica que a degradação ambiental nas áreas estudadas ainda não é acentuada, permitindo a migração das espécies litorâneas.
Dentre os Decapoda registradas para Angra dos Reis, as espécies Atya scabra, Macrobrachium carcinus, Potimirim glabra e P. potimirim estão classificadas como vulneráveis na lista das espécies ameaçadas de extinção no Estado do Rio de Janeiro. As espécies Macrobrachium acanthurus e Macrobrachium olfersii figuram na lista das espécies presemivelmente ameaçadas. Os principais fatores que ameaçam tais espécies são destruição do habitat, exploração predatória por caça, exploração por coleta, exploração por comércio etc.
Tabela 128 - Táxons de Crustacea ocorrentes na área de influência de Angra 3.
|
Bracuí |
P.Brava |
Mambucaba |
Coleções |
Atyidae |
|
|
|
|
Atya scabra |
|
|
|
X |
Potimirim glabra |
|
|
X |
X |
P. potimirim |
|
|
|
X |
Palaemonidae |
|
|
|
|
Macrobrachium acanthurus |
|
|
|
X |
M. carcinus |
|
|
|
X |
M. olfersii |
|
|
X |
X |
M. potiuna |
|
|
|
X |
Palaemon pandaliformes |
|
|
|
X |
Trychodactylidae |
|
|
|
|
Trichodactylus fluviatilis |
|
X |
|
X |
Insecta - Ephemeroptera
Considerando os resultados obtidos nas coletas e o material depositado em coleção (referente aos pontos controle), 24 táxons distribuídos em cinco famílias são registrados para a área, dos quais somente três espécies constavam na lista de dados existentes da literatura para o litoral sul do Estado do Rio de Janeiro (Tabela 129).
Tabela 129 - Táxons de Ephemeroptera ocorrentes na área de influência de Angra 3.
Clique aqui para visualizar a tabela 129
Provavelmente, maior esforço de coleta em outros sítios deve trazer registro de Campylocia bocainensis e Askola froehlichi. Cinco táxons somente ocorreram nos pontos controle. Dos 20 táxons coletados, 15 ocorreram também nos pontos controle. Por outro lado cinco espécies ocorreram pela primeira vez. A fauna de Ephemeroptera presente na área de estudos pode ser considerada rica em espécies e, devido ao incipiente conhecimento acercado grupo no Estado do Rio de Janeiro, há grande probabilidade de que novas espécies sejam encontradas.
Além dos registros novos para a área, o gênero Allenhyphes é ocorrência nova no Estado do Rio de Janeiro. A maioria dos táxons presentes é generalista em termos de qualidade de habitat, tais como as espécies de Cloeodes, Americabaetis, Caenis, Leptohyphes, Miroculis e Farrodes. Desdes dois últimos gêneros as espécies F. carioca e M. foehlichi ocorrem tanto em seções ritrais como em potamais de rios, incluíndo poças e represamentos (Da Silva, 2002) Algumas estão mais restritas a cursos d’água com maior correnteza e concentração de oxigênio, como Baetodes serratus, Camellobaetidius billi e Lepthohyphes inanis. Por outro lado, táxons de ampla distribuição e valência ecológica, como Massartella brieni e Americabaetis longetron, que provavelmente ocorrem na região, não foram encontrados.
Insecta - Odonata
No total, 67 espécies (morfótipos) de Odonata foram registradas no município de Angra dos Reis (Tabela 130). Os dados existentes, relacionados à listagem de Carvalho & Pujol-Luz (1992), para a Ilha Grande, reunidos àqueles das coleções estudadas, totalizaram 44 espécies. Nas coletas realizadas na área de influência da Usina Angra 3, foi coligido material referente a 36 espécies. Destas, 23 estão sendo registradas pela primeira vez para o município, o que indica o pouco conhecimento a respeito da ocupação do grupo no Estado do Rio de Janeiro.
O número total citado representa cerca de 25% das espécies registradas para o estado (Carvalho & Nessimian, 1998). Em relação ao Estado de São Paulo, apenas quatro espécies (Coryphaeschna adnexa, Mnesarete borchgravii, Metaleptobasis macilenta e Heteragrion consors) não constam da lista apresentada por Costa et al. (2000).
Tabela 130 - Táxons de Odonata ocorrentes na área de influência de Angra 3.
Clique aqui para visualizar a tabela 130
A composição de espécies encontrada é característica das áreas serranas da floresta atlântica, em sua maioria, representada por espécies de ampla distribuição no Sudeste do Brasil, sendo bastante similar àquela apresentada por Santos (1970) para o Parque Nacional de Itatiaia, RJ.
Pouco se sabe a respeito da biologia das espécies registradas, além da preferência dos sítios de criação dos imaturos da maioria. A espécie Mecistogaster asticta, registrada para o município, está indicada como “em perigo” na lista de espécies ameaçadas de extinção do Estado do Rio de Janeiro (Otero et al., 2000), em função de sua dependência a habitats itotelmatos umbrófilos. Esta espécie e Mnesarete borchgravii são provavelmente os odonatos mais sensíveis a alterações ambientais daqueles ocorrentes na área estudada, tendo sido registrados apenas nos dois pontos mais preservados, considerados como controles para os macroinvertebrados aquáticos (rio Caputera e rio Bonitinho).
Insecta - Plecoptera
Foram registrados para a área de estudos, com base nas coletas e em material depositado em coleções, seis dos oito gêneros ocorrentes no estado do Rio de Janeiro (Tabela 131). Contudo, nas coletas realizadas, somente Anacroneuria e Gripopteryx foram encontrados. A ordem está restrita a ambientes lóticos e águas limpas, frias e bem oxigenadas. A maioria dos grupos está relacionada a substratos rochosos e, no caso de Gripopteyix, à ocorrência de hidrófitas. Dentre os Perlidae, Anacroneuria é o gênero dominante em baixas altitudes e suas espécies são relativamente mais tolerantes a perturbações no habitat.
Tabela 131 - Táxons de Plecoptera ocorrentes na área de influência de Angra 3.
| |
Bracuí |
P.Brava |
Mambucaba |
Coleções |
| Perlidae |
|
|
|
|
| Anacroneuria sp. |
|
X |
X |
X |
| Kempnya sp. |
|
|
|
X |
| Macroginoplax sp. |
|
|
|
X |
| Gripopterygidae |
|
|
|
|
| Gripopteryx sp. |
|
X |
|
X |
| Paragrypopteryx sp. |
|
|
|
X |
| Tupiperla sp. |
|
|
|
X |
Insecta - Hemiptera
Foram registradas 71 espécies de Hemiptera (Tabela 132), incluindo morfótipos, na área de influência de Angra 3. Dessas espécies, 40 foram coletadas durante as visitas ao campo. Somente uma espécie registrada na bibliografia, Asterocoris australis, família Aradidae, com localidade-tipo em Angra dos Reis-RJ, foi relacionada em coleções. Esses dados revelam o pouco conhecimento acerca da fauna de hemípteros da região em questão.
Tabela 132 - Táxons de Hemiptera ocorrentes na área de influência de Angra 3.
Clique aqui para visualizar a tabela 132
Insecta - Homoptera
Foram registradas 163 espécies de homópteros (Tabela 133), incluindo morfótipos, na área de influência da Usina Angra 3. Nos trabalhos de campo de maio de 2002, foi coletado material referente a 89 espécies, o que corresponde a pouco mais da metade do total registrado. Tais espécies são relativamente comuns em áreas de Mata Atlântica, ocupando principalmente os ambientes de borda de mata. Algumas ocorrem em áreas mais abertas, estando relacionados com diferentes espécies de gramíneas. Nenhuma das espécies relacionadas na bibliografia para o município de Angra dos Reis (Zanol & de Menezes, 1982) consta no material observado.
Tabela 133 - Táxons de Homoptera ocorrentes na área de influência de Angra 3.
Com relação às coletas, foi verificada uma boa diversidade de espécies, considerando-se a estação do ano, final de outono, e o pequeno número de amostragens realizado. Esse fato é especialmente importante na tribo Proconiini, um grupo da família Cicadellidae. Membros desse táxon não são facilmente encontrados em campo e algumas espécies são pouco numerosas em coleções, como por exemplo Phera carbonaria, coletada na região de praia Brava. A diversidade de Membracidae também merece destaque. Por exemplo, foram coletadas sete espécies do gênero Cyphonia nas localidades estudadas. Vários exemplares de uma espécie ainda não descrita de Coronigoniella, família Cicadellidae, foram coletados em todas as grandes áreas visitadas.
Insecta - Megaloptera
Na área estudada, registrou-se exemplares imaturos de um único morfótipo de Corydalus (Corydalidea) (Tabela 134). Tal morfótipo foi encontrado apenas nas áreas Bracuí e Mambucaba.
Tabela 134 - Táxons de Megaloptera ocorrentes na área de influência de Angra 3.
| |
Bracuí |
P.Brava |
Mambucaba |
Coleções |
| Corydalidae |
|
|
|
|
| Corydalus |
X |
|
X |
X |
Insecta - Coleoptera
Registrou-se cerca de 155 espécies ou morfótipos de Coleoptera nas áreas de influência do empreendimento (Tabela 135). Das 90 espécies coletadas nas campanhas, 85 ainda não haviam sido registradas.
Tabela 135 - Táxons de Coleoptera ocorrentes na área de influência de Angra 3.
Clique aqui para visualizar a tabela 135
Dentre os Insecta considerados neste estudo, os Coleoptera são extremamente pouco conhecidos na região de Angra dos Reis, devendo chegar a alguns milhares de espécies. Zikán & Wygodzinsky (1948) relacionaram apenas duas espécies para o município de Angra dos Reis. Correspondendo à maior ordem de Insecta, os Coleoptera ainda não foram alvo de estudados sistematicos na região, sendo os dados existentes considerados insuficientes para qualquer abordagem. A falta de conhecimento acumulado a respeito do grupo só poderá ser resolvida com coletas sucessivas por vários anos e com o trabalho continuado de especialistas das mais diversas famílias.
No presente estudo, a família Chrysomelidae foi a que apresentou o mais número de espécies, com cerca de 50, e a mais freqüentemente encontrada. Algumas espécies são relativamente comuns em áreas de Mata Atlântica ou áreas mais abertas, estando relacionadas com diferentes espécies vegetais. Dentre o material de coleópteros aquáticos estudado, foram encontrados exemplares de duas espécies de Elmidae ainda não descritas.
Insecta - Trichoptera
Foram registrados para a área de influência da Usina Angra 3, com base nas coletas realizadas e material depositado em coleções (referente aos pontos controle), 38 táxons de Trichoptera, entre espécies e morfótipos. (Tabela 136). Desses, 32 táxons ocorreram nas coletas.
Tabela 136 - Táxons de Trichoptera ocorrentes na área de influência de Angra 3.
Clique aqui para visualizar a tabela 136
Com relação aos dados registrados anteriormente por Carrijo et al. (2001), não foram encontrados representantes dos gêneros Mortoniella, Scactobiella, Barypenthus e Polycentropus (embora exemplares da família Polycentropodidae, não identificados, tenham sido coletados). Os exemplares de Leptonema e Macronema não foram identificados no nível de espécie, o que impede a confirmação da presença de L. pallidum e M. fulvum na área de estudo. Por outro lado, 12 táxons são registrados pela primeira vez: Contulma, Mexitrichia, Dolophiloides (Sortosa), Cernotina, Polyplectropus, Xiphocentron e mais seis espécies/morfoespécies de Chimarra. Mexitrichia é registro novo para o Estado do Rio de Janeiro.
A maioria dos grupos presentes é típica da seção ritral de rios, estando associada a substrato rochoso e respondendo negativamente a baixas concentrações de oxigênio. Dentre os táxons mais comumente encontrados e de maior distribuição, estão Leptonema, Smicridea, Triplectides e Chimarra. Contudo, a riqueza de táxons da ordem na área pode ser considerada alta, o que reflete habitats diversificados e em bom estado de conservação.
Insecta - Lepidóptera
Considerou-se, para este relatório, somente o grupo dos Rhopalocera (lepidópteros diurnos), por ser o melhor conhecido dessa ordem. Foram catalogadas 588 espécies de borboletas na área de influência da Usina Angra 3 (Anexo 13).
Somente 88 foram coletadas nas visitas ao campo, sendo que 13 dessas espécies não foram assinaladas nos dados provenientes de coleções, bem como nos dados bibliográficos, podendo constituir novos registros.
Como aspecto relevante, a região em questão faz parte da área de distribuição geográfica conhecida original de quatro espécies ameaçadas de extinção: Arcas ducalis, Agrias claudina, Heraclides himeros e Parides ascanius (Otero et al., 2000). As populações de tais espécies estão aparentemente restritas a pequenas áreas no Estado do Rio de Janeiro, devido à degradação de seus ambientes naturais (Otero et al., 2000).
Insecta - Diptera
Até o momento, 80 morfótipos de Diptera foram registradas no município de Angra dos Reis (Tabela 137). Como citado anteriormente, os Diptera, assim como os Coleoptera, provavelmente são os menos conhecidos na região em questão, devendo existir alguns milhares de espécies.
Tabela 137 - Táxons de Diptera ocorrentes na área de influência de Angra 3.
Clique aqui para visualizar a tabela 137
Algumas das maiores famílias em número de espécies nunca foram estudadas sistematicamente na região, sendo os dados existentes considerados insuficientes para qualquer abordagem. Espécies ruderais invasoras como Musca domestica e Chrysomyia spp. sequer haviam sido registradas para a região. Poucos artigos, como o de Zikán & Wygodzinsky (1948) e Lopes (1973), listam espécies de Stratiomyiidae, Ephydridae e Sarcophagidae para o município de Angra dos Reis. As duas listagens produzidas no âmbito deste estudo apresentam poucos morfótipos em comum. Tal efeito se refere à falta de conhecimento acumulado a respeito do grupo, ao nível pouco apurado de identificação e ao número reduzido de coletas realizadas. Esses problemas só poderão ser resolvidos com coletas sucessivas por vários anos e com o trabalho continuado de especialistas das mais diversas famílias (inclusive estrangeiros).
Embora uma lista dos Culicidae (mosquitos) de uma região vizinha ao município de Angra dos Reis tenha sido incluída no levantamento pretérito (Guimarães et al., 2000), durante as coletas realizadas em maio de 2002 esse grupo não foi enfocado.
Macroinvertebrados aquáticos
Considerando os dados provenientes das coletas realizadas nos pontos amostrais e de material dos pontos controle (14 e 15), os macroinvertebrados aquáticos somaram 190 táxons de Insecta (187) e Crustacea Decapoda (3). Destes, 114 ocorreram na amostragem realizada. Contudo, em termos de riqueza taxonômica por ponto de coleta, devido, além de outros fatores, ao método e ao esforço empregados, há uma grande diferença nos resultados. Os valores obtidos nos pontos controle, nos quais o esforço foi maior, foram 116 e 122, enquanto o valor máximo dentre os pontos amostrais foi 41, no rio Água Branca. Embora o peso relativo à ordem Diptera (especialmente Chironomidae, cuja acuidade taxonômica varia com o método de coleta) seja alto, as diferenças seriam significativas mesmo com sua retirada dos cálculos (81 e 77 contra 34) (Figura 142).
Figura 142 - Número de táxons de macroinvertebrados aquáticos obtidos nos trechos de rio onde foram realizadas as amostragens de macroinvertebrados aquáticos e nos rios controle
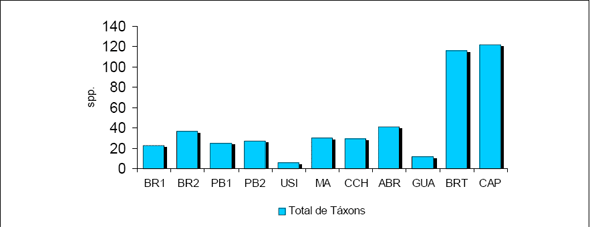
Obs.: ABR - Rio Água Branca; BRP - Rio Bracuí (seção potamal); BRR - Rio Bracuí (seção ritral); BRT - Tributário do Rio Bracuí, CAP - Rio Caputera; CCH - Rio Cachoeirinha; GUA - Rio Guariri; MA - Rio Mambucaba (seção potamal); PB1 - Tributário do Rio Cachoeira Brava; PB2 - Rio Cachoeira Brava; USI - Riachos próximos à planta da Usina Nuclear.
O número de táxons EPT (Ephemeroptera, Plecoptera e Trichoptera) constitui uma boa medida bioindicadora da integridade de rios (Rosemberg & Resh, 1993). Comparando-se os rios amostrados e os rios controle, nota-se também grande diferença. Nos pontos controle o número de EPT variou entre 39 e 42 enquanto o valor máximo obtido nos pontos amostrais foi 21. Provavelmente, maior esforço de amostragem será necessário. Contudo, notam-se diferenças significativas entre os pontos amostrais, com valores mais baixos para o Rio Guariri e a seção potamal do rio Bracuí e nenhum táxon encontrato nos córregos próximos à Angra 3 (Figura 143).
Figura 143 - Número de táxons EPT (Ephemeroptera, Plecoptera e Trichoptera) obtidos.
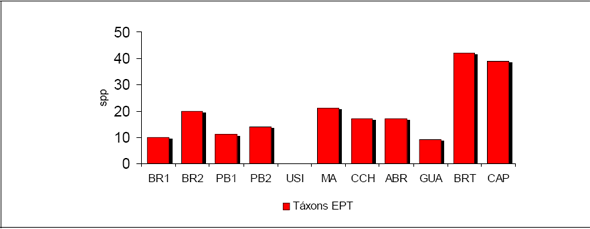
Obs.: ABR - Rio Água Branca; BRP - Rio Bracuí (seção potamal); BRR - Rio Bracuí (seção ritral); BRT - Tributário do Rio Bracuí, CAP - Rio Caputera; CCH - Rio Cachoeirinha; GUA - Rio Guariri; MA - Rio Mambucaba (seção potamal); PB1 - Tributário do Rio Cachoeira Brava; PB2 - Rio Cachoeira Brava; USI - Riachos próximos à planta da Usina Nuclear.
Outra medida utilizada para avaliar a integridade dos rios foi o BMWP adaptado de Junqueira et al. (1998), no qual são atribuidos valores numéricos para cada família presente de acordo com seu grau de tolerância. Os valores mais baixos corresponderam aos córregos próximos à Usina. Mais uma vez, os rios controle apresentaram valores significativamente mais altos. Os rios controle podem ser considerados em estado “excelente”, os rios Cachoeira Brava e Água Branca, “muito bom”, os córregos próximos à usina, “pobre” e os demais, “bom”.
Na Figura 144 são apresentados os valores obtidos de BMWP e do protocolo de avaliação fisionômica de Petersen (1992) para cada ponto de coleta. Os valores mais baixos correspondem aos córregos próximos à Usina e à seção potamal do Rio Bracuí. Como o número de famílias presentes tem relação direta com os valores de BMWP, os valores obtidos foram divididos pelo número de famílias (BMWP-ASTP).
Figura 144 - Valores obtidos na avaliação fisionomica (Petersen, 1992) e BMWP (adaptado de Junqueira et al. 1998).
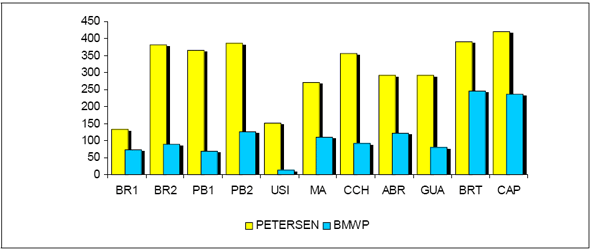
Obs.: ABR - Rio Água Branca; BRP - Rio Bracuí (seção potamal); BRR - Rio Bracuí (seção ritral); BRT - Tributário do Rio Bracuí, CAP - Rio Caputera; CCH - Rio Cachoeirinha; GUA - Rio Guariri; MA - Rio Mambucaba (seção potamal); PB1 - Tributário do Rio Cachoeira Brava; PB2 - Rio Cachoeira Brava; USI - Riachos próximos à planta da Usina Nuclear).
Nota-se mais uma vez o menor valor nos córregos próximos à usina. Contudo, para aos outros pontos de amostragem e para os pontos controle, os valores foram similares.
Figura 145 - Valores obtidos de BMWP-ASTP (BMWP dividido pelo número de famílias presentes)
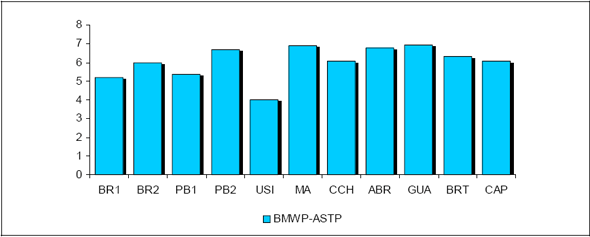
Obs.: ABR - Rio Água Branca; BRP - Rio Bracuí (seção potamal); BRR - Rio Bracuí (seção ritral); BRT - Tributário do Rio Bracuí, CAP - Rio Caputera; CCH - Rio Cachoeirinha; GUA - Rio Guariri; MA - Rio Mambucaba (seção potamal); PB1 - Tributário do Rio Cachoeira Brava; PB2 - Rio Cachoeira Brava; USI - Riachos próximos à planta da Usina Nuclear.
Com relação à composição de táxons, a fauna encontrada reflete o tipo (lêntico ou lótico, ritral ou potamal), o tamanho e o estado de preservação dos corpos d’água. Como assinalado anteriormente na apresentação das ordens consideradas para este levantamento, táxons de distribuição restrita ou com maiores exigências quanto ao habitat, ocorreram somente em locais com maoires valores de integridade. Nas tabelas são apresentados os resultados da distribuição dos táxons de macroinvertebrados aquáticos nos pontos de coleta.
Tabela 138 - Taxons de Decapoda (Crustacea) ocorrentes na área de influência de Angra 3.(Material coletado em corpos d’água nas três áreas estabelecidas e nos pontos controle (CONTR.)).
Clique aqui para visualizar a tabela 138
Tabela 139 - Táxons de Ephemeroptera ocorrentes na área de influência de Angra 3. (Material coletado em corpos d’água nas três áreas estabelecidas e nos pontos controle (CONTR.)).
Clique aqui para visualizar a tabela 139
Tabela 140 - Táxons de Odonata ocorrentes na área de influência de Angra 3 (Material coletado em corpos d’água nas três áreas estabelecidas e nos pontos controle (CONTR.)).
Clique aqui para visualizar a tabela 140
Tabela 141 - Táxons de Plecoptera ocorrentes na área de influência de Angra 3 (Material coletado em corpos d’água nas três áreas estabelecidas e nos pontos controle (CONTR.)).
Clique aqui para visualizar a tabela 141
Tabela 142 - Táxons de Hemiptera aquáticos ocorrentes na área de influência de Angra 3 (Material coletado em corpos d’água nas três áreas estabelecidas e nos pontos controle (CONTR.)).
Clique aqui para visualizar a tabela 142
Tabela 143 - Táxons de Megaloptera ocorrentes na área de influência de Angra 3 (Material coletado em corpos d’água nas três áreas estabelecidas e nos pontos controle (CONTR.)).
Clique aqui para visualizar a tabela 143
Tabela 144 - Taxons de Coleoptera aquáticos ocorrentes na área de influência de Angra 3 (Material coletado em corpos d’água nas três áreas estabelecidas e nos pontos controle (CONTR.))
Clique aqui para visualizar a tabela 144
Tabela 145 - Táxons de Trichoptera ocorrentes na área de influência de Angra 3 (Material coletado em corpos d’água nas três áreas estabelecidas e nos pontos controle (CONTR.)).
Clique aqui para visualizar a tabela 145
Tabela 146 - Táxons de Diptera aquáticos ocorrentes na área de influência de Angra 3 (Material coletado em corpos d’água nas três áreas estabelecidas e nos pontos controle (CONTR.))
Clique aqui para visualizar a tabela 146
O número de táxons de Insecta registrados para a região de Angra dos Reis (cerca de 1200) não reflete, ainda, a magnitude da diversidade esperada para aréas de Mata Atlântica. Contudo, foram bastante representativos os dados obtidos para alguns grupos, como Lepidoptera diurnos e Odonata. Do material coletado, 272 táxons representam novos registros para a região de Angra dos Reis.
Para o Estado do Rio de Janeiro, os gêneros Allenhyphes (Ephemeroptera) e Mexitrichia (Trichoptera) foram registrados pela primeira vez. A partir das coletas realizadas, há a provavel ocorrência de várias espécies ainda não descritas para a ciência, das quais três já foram confirmadas, nas ordens Homoptera e Coleoptera.
Foi constatado que a região estudada faz parte da área de distribuição geográfica de algumas espécies incluídas na “Lista de espécies ameaçadas de extinção do Estado do Rio de Janeiro” (Bergallo et al., 2000). As espécies Agrias claudina, Heraclides himeros e Parides ascanius (Insecta, Lepidoptera) estão classificadas como “criticamente em perigo”; Mecistogaster asticta (Insecta, Odonata), com “em perigo”; e Arcas ducalis (Insecta, Lepidoptera), Atya scabra, Macrobrachium carcinus, Potimirim glabra e P. potimirim (Crustacea, Decapoda), como “vulneráveis”. As espécies Macrobrachium acanthurus e Macrobrachium olfersii (Crustacea, Decapoda) figuram na lista das espécies presumivelmente ameaçadas. O principal fator que ameaça tais espécies é a destruição de seus habitats naturais.
Grande parte dos táxons coletados na área de influência de Angra 3 apresenta-se relacionada a ambientes alterados e periurbanos, com a presença também de espécies exóticas. São exemplos, Pantala flavescens, Tramea cophysa, Orthemis discolor, Erithrodiplax umbrata, E. fusca (Odonata), Dryas julia, Eurema spp., Anteos menipe, Ascia monuste (Lepidoptera), Musca domestica, Chrysomyia spp. (Diptera), Deois spp., Versigonalia ruficauda, Sibovia sagata (Homoptera), Dysdercus sp. (Hemiptera) e várias espécies de Chrysomelidae (Coleoptera)
A fauna de macroinvertebrados aquáticos apresentou-se mais pobre em táxons em corpo d’água de áreas mais perturbadas ou alteradas. Dentre os macroinvertebrados aquáticos, táxons com maior restrição com relação a habitats e menos tolerantes ocorreram somente nos locais menos alterados, como por exemplo, Baetodes serratus, Camellobaetidius billi e Lepthohyphes inanis (Ephemeroptera), Mnesarete borchgravii, Heteragrion spp. (Odonata), Kempnyia spp. e Paragripopteryx spp. Gripopteryx spp. (Plecoptera).
7.6. Incrustação - topo
O termo incrustação biológica ("fouling") é usualmente empregado para distinguir os grupos de animais e plantas que crescem em substratos artificiais imersos na água do mar, daqueles que ocorrem em costões, pedras ou outros substratos duros naturais. Este conceito é, portanto, um conceito prático, advindo da importância atribuída a estes organismos e seus efeitos, na sua maioria danosos, às estruturas construídas pelo homem, não havendo uma distinção biológica entre essas comunidades e as dos substratos naturais (WHOI,1952). Os grupos encontrados são predominantemente sésseis e representados principalmente por cirripédios, ascídias, hidrozoários, briozoários e algas, seguidos de bivalves, esponjas e corais.
A composição da comunidade incrustante encontrada num determinado substrato artificial imerso vai depender das espécies naturalmente presentes naquela localidade e de suas habilidades para se fixar e crescer nessas superfícies. Além disso, as estratégias reprodutivas das diferentes espécies ali presentes irão determinar que organismos colonizarão o substrato nas diferentes épocas do ano (Sutherland, 1974; Sutherland & Karlson, 1977).
O estabelecimento de uma nova comunidade incrustante, a partir de uma superfície "desnuda", dá-se a partir de um tipo de sucessão ou seqüência temporal de táxons. Organismos microscópicos (bactérias, diatomáceas, cianofíceas e protozoários) são os primeiros a colonizarem o substrato recém submerso, multiplicando-se rapidamente (Keough & Raimondi, 1995). Posteriormente, aparecem as formas macroscópicas, cujas larvas estão presentes na época de imersão do substrato (Valiela, 1984). Após a colonização inicial, o aumento da biomassa depende basicamente da taxa de crescimento dos organismos assentados, fato que difere de espécie para espécie e é controlado, principalmente, pela temperatura da água e disponibilidade de alimento. Ao longo do tempo, ocorre um aumento do número de táxons assentados, ocorrendo variações de riqueza entre as diferentes épocas do ano.
No interior dos circuitos de refrigeração de usinas nucleares que utilizam a água do mar, como a CNAAA, ocorre o desenvolvimento de incrustação, e esta colonização biológica diminui a eficiência na troca de calor e gera problemas de corrosão nas paredes das tubulações. Deste modo, a avaliação da bioincrustação na área da tomada d'água permite identificar as principais espécies presentes naquele local e seus períodos de recrutamento, o que pode vir a aprimorar o sistema de cloração utilizado em cada central nuclear a cada época de ano; controlando efetivamente os efeitos negativos da incrustação no interior das tubulações (Cole, 1977). A avaliação das comunidades incrustantes em painéis artificiais é extremamente útil (Richmond & Seed, 1991), e auxilia tanto no controle eficaz da incrustação no interior das usinas quanto para avaliar o impacto destas usinas nos corpos receptores da água de refrigeração. Desta forma, o uso de painéis artificiais também é uma estratégia eficaz para o monitoramento das áreas sob influência do efluente de refrigeração, permitindo avaliar seus impactos sobre a biota e verificar até onde essa interferência é observada.
São escassos os trabalhos realizados com comunidades incrustantes na Baía de Ilha Grande e, dos 10 trabalhos analisados, apenas 6 apresentam uma lista detalhada de espécies.
O trabalho mais antigo sobre comunidades incrustantes na Baía da Ilha Grande (Nunes, 1983) é um relatório técnico sobre o “fouling” nas imediações da Central Nuclear. O estudo compara a incrustação biológica em dois tipos de substrato (madeira e amianto) nas fases pré e pós operacional da Unidade 1 da CNAAA (Angra 1). Foi constatado aumento de biomassa na descarga do efluente na fase pós operacional em 3 períodos do ano (outono, inverno e primavera). Cabe ressaltar que a temperatura da descarga na fase pós operacional não superou 27 °C.
Os trabalhos de Silva (1985) e Silva et al.(1989), foram os primeiros a analisar a relação entre a intensidade da incrustação biológica e a abundância de organismos perfurantes de madeira. Nestes trabalhos a abordagem em relação às comunidades incrustantes é apenas quantitativa, considerando variações temporais na biomassa total.
Roiff (1996) trabalhou em uma escala espacial maior, abrangendo 8 diferentes áreas da região sudeste (de São Sebastião até Vitória incluindo Angra dos Reis). Embora tenha verificado uma redução da biomassa total em direção ao Norte, conclui-se que a abundância e composição específica em cada estação de coleta deveu-se mais aos fatores locais como poluição orgânica, turbidez e pluviosidade do que a efeitos latitudinais de temperatura.
Skinner (1997) avaliou o efeito da salinidade sobre a composição de comunidades incrustantes ao longo do Rio Bracuhy e em estações externas ao rio.
Silva (1998) registrou a ocorrência de aproximadamente 85 táxons incrustantes em painéis de PVC, distribuídos em 3 estações sob diferentes graus de impacto orgânico. As comunidades na região sob maior impacto apresentaram o desenvolvimento mais rápido e a menor riqueza e diversidade de espécies dentre as 3 regiões estudadas. A região moderadamente impactada apresentou a maior riqueza e diversidade de espécies, porém com uma velocidade de desenvolvimento intermediária. A região não impactada apresentou o desenvolvimento mais lento e uma riqueza e diversidade um pouco menor que a região moderadamente impactada.
Falcão (1999) volta a analisar a relação entre a interferência da incrustação sobre os perfurantes verificando se esta ocorre na fase de pré-fixação das larvas ou sobre os adultos instalados nas galerias. O trabalho aborda a incrustação biológica por grupos funcionais e sua porcentagem de cobertura e avalia a biomassa total de cada unidade amostral.
Mayer (2001), dando continuidade ao trabalho de Silva (1998), realizou transferências de painéis artificiais entre uma área impactada e uma não impactada com o objetivo de avaliar o efeito da eutrofização no desenvolvimento das comunidades incrustantes. A eutrofização alterou a estrutura das comunidades apenas em relação à abundância das diferentes espécies, mas não alterou a sua composição específica original. O grupo que melhor refletiu as diferenças entre as áreas foi aquele representado por tubos mucosos de poliquetas e anfípodas.
Em relação ao impacto do efluente de refrigeração da CNAAA, foram realizadas recentemente duas pesquisas (Viana, 2001; Curbelo Fernandez, 2002) que utilizaram as comunidades incrustantes em painéis de madeira na avaliação deste impacto. O impacto da descarga do efluente provocou uma menor diversidade de espécies nas estações sob influência direta do fluxo da descarga e nas demais estações localizadas na área de descarga. Estes resultados foram observados pelas duas autoras, tanto para painéis mensais (colonização inicial) quanto para os painéis trimestrais (comunidade desenvolvida). Nos painéis mensais, durante 6 meses de estudo, foram registrados 32 táxons de macroorganismos. Nos trimestrais, durante 1 ano, foram registrados 52 táxons. Houve uma acentuada diminuição da diversidade da comunidade incrustante nas estações impactadas, principalmente, nos meses de primavera e verão, o que não foi verificado nas estações controle. Isto demonstra os efeitos danosos relacionados, principalmente, à elevada temperatura da água do efluente de refrigeração, não descartando os efeitos negativos do fluxo elevado e do cloro.
Os trabalhos acima confirmam a utilidade das comunidades incrustantes na avaliação de impactos ambientais, refletindo em sua estrutura e desenvolvimento as condições imperantes no ambiente. Além disso, demonstram que um grande número de táxons ocorre nestes substratos artificiais, refletindo de forma adequada a comunidade local e o recrutamento de larvas de organismos bentônicos dos substratos consolidados.
Devido as condições metereológicas desfavoráveis, várias unidades experimentais foram perdidas; e como o período para a avaliação deste estudo foi curto, tornou-se impossível recomeçar os trabalhos, substituindo-se as unidades experimentais.
Os trabalhos de Viana (2001) e Curbelo Fernandez (2002), na mesma área deste estudo, mostraram que a temperatura entre as estações da região controle e as da região impactada são distintas durante todo o ano, com ΔT aproximado de 8ºC para o período de abril a julho de 2001. No presente trabalho, valores similares foram observados. Além do impacto térmico, as estações sob efeito da descarga do efluente estão sujeitas aos efeitos nocivos do cloro e do fluxo elevado do efluente, sendo as três formas de impacto maiores quanto mais próxima a estação se encontra da área da descarga.
Para o estudo de incrustação foram estabelecidas seis estações experimentais no Saco Piraquara de Fora (descarga do efluente de refrigeração) e três em locais controle (Ilha do Brandão e Itaorna), conforme indicado na Figura 146. A Tabela 147 apresenta os nomes atribuídos a cada estação, sendo que a localizada na saída do efluente da usina, no Saco Piraquara de Fora, foi considerada como ponto de referência para o estabelecimento das demais.
Como corpo de prova para a bioincrustação foram utilizados painéis de granito de 20 x 20 x 2 cm (três réplicas por estação). Para as estruturas que permaneceram imersas na coluna d’água, foi traçada uma radial acompanhando a pluma térmica gerada pelo efluente da CNAAA nas estações Descarga, 600m/sentido Ilha Pingo d’Água e 1400m, no Saco Piraraquara de Fora, Controle 1, na praia de Itaorna, e Controle 2, na Ilha do Brandão. Nos costões rochosos, os granitos permaneceram fixos à rocha por cabos de aço e cimento (tubolite), abaixo da linha mínima de maré baixa nas estações 600m/sentido Ilha Pingo d’água, 600m/sentido Ponta Grossa e 1400m, no Saco Piraquara de Fora; e no Controle 2, na Ilha do Brandão. Em todos os casos, os painéis permaneceram submersos a uma profundidade de 80 cm.
Tanto as estruturas que permaneceram fixas no costão quanto as submersas na coluna d’água foram colocadas no dia 19 de abril e retiradas em 7 de julho de 2002, totalizando 80 dias de imersão. Depois de retirados, os painéis foram acondicionados em sacos plásticos e congelados a -20°C.
Figura 146 - Incrustação - Estações experimentais (em vermelho,costão, e em azul, coluna d’água).
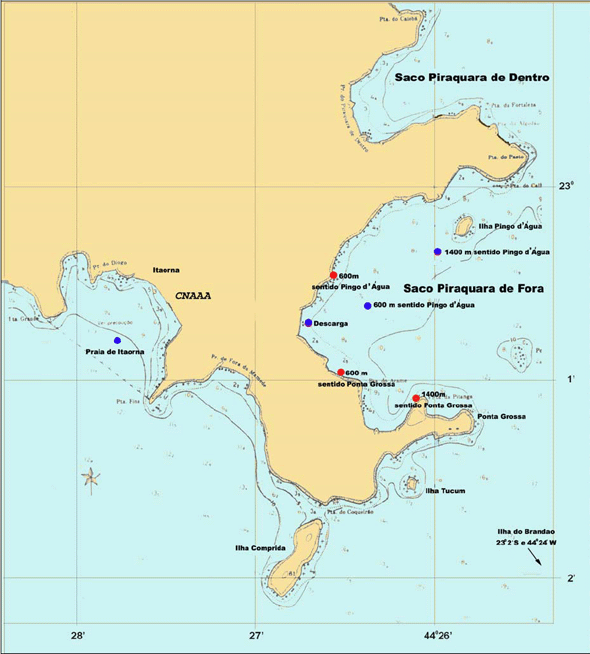
Tabela 147 - Incrustação - Estações experimentais
Região |
Experimento |
Local |
Estação |
Área impactada |
Costão |
Piraquara de Fora, sentido Ilha Pingo d’Água |
600m |
| Piraquara de Fora, sentido Ponta Grossa |
600m |
| Piraquara de Fora, sentido Ponta Grossa |
1.400m |
Coluna d’água |
Piraquara de Fora, próximo à saída do efluente |
Descarga |
| Piraquara de Fora, sentido Ilha Pingo d’Água |
600m |
| Piraquara de Fora, sentido Ilha Pingo d’Água |
1400m |
Áreas de controle |
Costão |
Ilha do Brandão |
Controle 2 |
| Coluna d’água |
Ilha do Brandão |
Controle 2 |
| Praia da Itaorna |
Controle 1 |
Em laboratório, para a análise quantitativa, os painéis foram descongelados e uma de suas faces foi analisada sob microscópio estereoscópico, sendo os organismos incrustantes identificados e contados pelo método de pontos de interseção descrito por Sutherland (1974). A análise estatística (análise de variância e testes t) foi realizada considerando apenas as espécies registradas por esse método.Complementarmente, foram registradas as espécies presentes nos painéis mas não identificadas pelo método de pontos de interseção.
Considerando todos os painéis experimentais (bóias e costões) foi observada ocorrência nova de 12 espécies de macroalgas para a região de estudo.
Bóias
A ausência de réplica para os painéis, de modo a permitir um tratamento estatístico adequado dos dados, levou a uma análise qualitativa deste tratamento.
De um modo geral, os painéis que permaneceram submersos na coluna da água, apresentaram maior cobertura de invertebrados do que os painéis fixos ao costão. Além de 6 espécies de cirripédios, estes painéis apresentaram briozoários incrustantes (2 espécies), ascídia colonial (1 espécie) e bivalve (1 espécie). As macroalgas apresentaram menores coberturas e riqueza quando comparadas com os painéis do costão. A lista completa da presença de espécies nestes painéis, encontra-se na Tabela 148.
O impacto da descarga do efluente da CNAAA sobre a comunidade incrustante se refletiu na riqueza de táxons registrada nos painéis ao longo da radial. A Tabela 149 mostra a riqueza de táxons para os painéis artificiais utilizando a metodologia de pontos de interseção e a riqueza total, que considerou também a avaliação qualitativa dos painéis.
No Saco Piraquara de Fora, a estação mais próxima da saída do efluente de refrigeração (Descarga) apresentou a menor riqueza de táxons da área. Nas bóias localizadas a 600m da saída, o número de táxons aumentou sensivelmente, para atingir o máximo na estação localizada a 1400m da descarga do efluente da CNAAA. Nas estações Controle 1 (Praia de Itaorna) e Controle 2 a riqueza apresentou valores próximos aos encontrados na estação 1400 m sentido Pingo D`água.
Tabela 148 - Lista dos táxons presentes para o tratamento coluna d’água (bóias), para as estações analisadas.
Clique aqui para visualizar a tabela 148
Tabela 149 - Riqueza de táxons registrados nos painéis artificiais submersos nas 5 estações experimentais.
Clique aqui para visualizar a tabela 149
Costão
Para este tratamento foi possível uma análise estatística adequada dos dados, e a lista completa de espécies de macroalgas e invertebrados bentônicos encontrados nestes painéis, encontra-se na Tabela 150.
Tabela 150 - Lista dos táxons presentes para o tratamento costão, para as estações analisadas
Clique aqui para visualizar a tabela 150
O biofilme, composto por algas cianofíceas, diatomáceas, protozoários, bactérias, matéria orgânica e inorgânica, não apresentou diferenças significativas na cobertura entre as estações da área impactada e as estações das áreas controle (Figura 147).
Figura 147 - Porcentagem de cobertura do Biofilme (média e erro padrão) para as 4 estações analisadas.
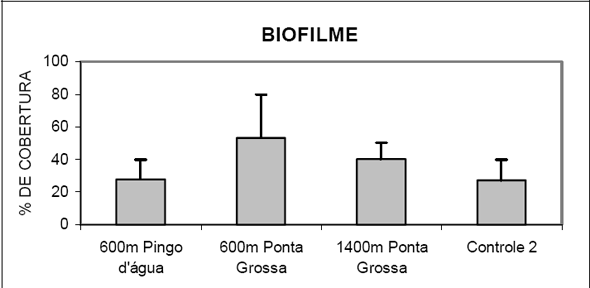
No entanto, a porcentagem de cobertura do biofilme nas estações da área sob influência da CNAAA foi, em alguns casos, superior a 50 % (média da estação), enquanto que na estação Controle 2 na Ilha do Brandão, a cobertura média foi inferior a 30%. O aumento na porcentagem de cobertura do biofilme em locais sujeitos ao impacto do efluente aquecido de uma usina nuclear foi observado, também, por outros autores na Flórida (Thorhaug et al., 1971; Roessler, 1971). Em relação às macroalgas, a cobertura de Chlorophyta foi significativamente superior na estação Controle 2 quando comparada com a estação 600m sentido Ponta Grossa (próxima à descarga do efluente) (Figura 148). Em todas as estações, os gêneros Cladophora e Enteromorpha foram os mais abundantes.
Figura 148 - Porcentagem de cobertura de Chlorophyta (média e erro padrão) para as 4 estações analisadas.
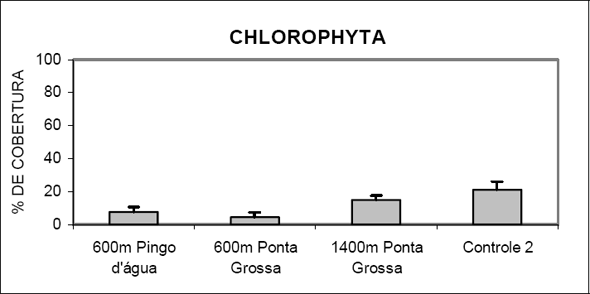
As algas da divisão Rhodophyta foram as mais abundantes neste estudo, e não apresentaram diferenças significativas na porcentagem de cobertura entre as 4 estações analisadas (Figura 149).
Figura 149 - Porcentagem de cobertura de Rhodophyta (média + erro padrão) para as 4 estações analisadas.
É importante observar que a espécie predominante em cada uma das estações foi diferente. Na estação 600m Pingo D’água a espécie predominante foi Acanthophora spicifera; para 600m sentido Ponta Grossa, Polysiphonia subtilissima foi a espécie predominante; já para 1400m sentido Ponta Grossa (a mais afastada da saída do efluente dentro do Saco Piraquara de Fora) a espécie foi Jania Capillacea; e para a estação Controle 2 (Ilha do Brandão) a espécie predominante foi Laurencia papillosa. As macroalgas da divisão Phaeophyta não apresentaram diferenças significativas na porcentagem de cobertura entre as estações a 1400m sentido Ponta Grossa e Controle 2 (Ilha do Brandão) (Figura 150).
Figura 150 - Porcentagem de cobertura de Phaeophyta (média + erro padrão) para as 4 estações analisadas.
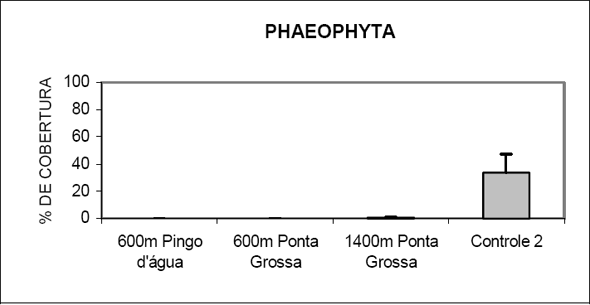
Embora não tenha sido observada diferença significativa entre a cobertura da divisão Phaeophyta entre as duas estações onde este grupo ocorreu (1400 m Ponta Grossa e Ilha do Brandão - controle 2), a estação Controle 2 apresentou maior porcentagem de cobertura em todas as réplicas analisadas. As espécies mais abundantes nesta estação foram Ralfsia expansa e Hincksia mitchelliae. Esta divisão (Phaeophyta) parece ser a mais afetada pelas altas temperaturas do efluente, já que só foi observada nas estações mais afastadas da descarga. Com relação a essas algas, Curbelo Fernandez (2002) encontrou resultados similares aos nossos.
Os únicos invertebrados marinhos registrados nos painéis fixados aos costões foram 2 espécies de crustáceos cirripédios encontrados em uma das estações do Saco Piraquara de Fora (600m sentido Ponta Grossa) e na estação Controle 2 (Ilha do Brandão). Duas espécies de briozoários incrustantes foram registradas unicamente na estação Controle 2 (Ilha do Brandão) e com baixa porcentagem de cobertura (<5%).
Da mesma maneira que para os painéis experimentais imersos na coluna d’água, os painéis fixos ao costão apresentaram as menores riquezas de táxons nas estações mais próximas à descarga do efluente da CNAAA (Tabela 151). Dentro do Saco Piraquara de Fora, a estação mais afastada da saída do efluente (1400m sentido Ponta Grossa), apresentou um aumento do número de táxons registrados, indicando, provavelmente, uma diminuição do impacto do efluente a esta distância da descarga. Mais uma vez, a estação Controle 2 (Ilha do Brandão) foi a que apresentou a maior riqueza.
Tabela 151 - Riqueza de táxons registrados nos painéis artificiais fixados no costão nas 4 estações experimentais .
| |
600 m sentido Pingo D`água |
600 m sentido Ponta Grossa |
1400 m sentido Ponta Grossa |
Ilha do Brandão (controle 2) |
| Quantitativo |
2 |
6 |
8 |
16 |
| Total |
10 |
6 |
15 |
18 |
Painéis mensais
Os resultados apresentados a seguir correspondem a um trabalho paralelo desenvolvido pelo Laboratório de Benthos na área de influência da CNAAA. Os painéis permaneceram imersos na coluna de água a uma profundidade aproximada de 0,5 m durante um mês dentro do período do presente estudo, em estruturas similares às utilizadas no caso dos painéis trimestrais. As estações consideradas neste estudo foram: Descarga e 600m sentido Ponta Grossa (estações sob impacto) e os Controles 1 e 2.
Em relação ao biofilme (Figura 151), a cobertura foi significativamente superior na estação Descarga, com valores de até 100%. Já nas estações controle, a cobertura máxima do biofilme foi, aproximadamente, de 50%. Estes dados estão de acordo com o anteriormente apresentado para painéis trimestrais.
Figura 151 - Porcentagem de cobertura de biofilme (média + erro padrão) para as 4 estações analisadas no presente trabalho.
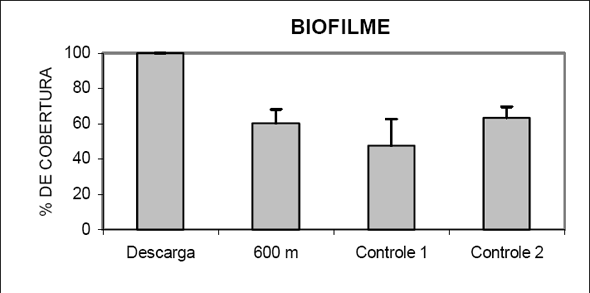
Para as macroalgas da divisão Chlorophyta (Figura 152), a porcentagem de cobertura foi significativamente superior na estação 600 m sentido Ponta Grossa (30 % de cobertura).
Figura 152 - Porcentagem de cobertura de Chlorophyta (média + erro padrão) para as 4 estações analisadas.
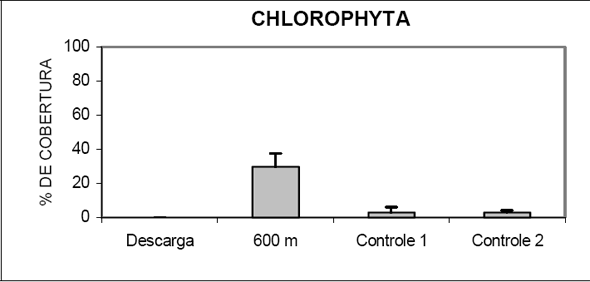
A divisão Rhodophyta, por sua vez, não apresentou diferenças significativas entre as estações analisadas (Figura 153), embora as maiores porcentagens de cobertura tenham correspondido à estação Controle 1 (Praia de Itaorna).
Figura 153 - Porcentagem de cobertura de Rhodophyta (média + erro padrão) para as 4 estações analisadas.
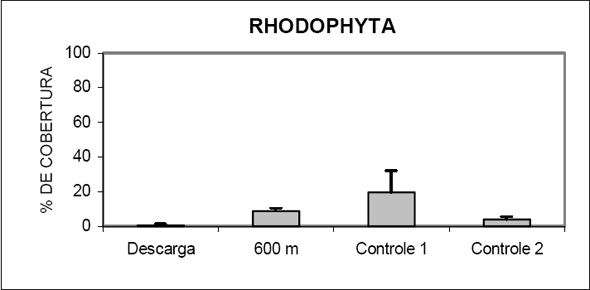
O grupo Hydrozoa só foi observado nas duas estações controle, e apresentou coberturas aproximadas de 30% em ambas as estações e sem diferenças significativas entre elas (Figura 154). A ausência deste grupo na área sob impacto já foi observada por Viana (2001) e Curbelo Fernandez (2002), relacionando a ausência do grupo Hydrozoa ao impacto da temperatura elevada e do fluxo.
Figura 154 - Porcentagem de cobertura de Hydrozoa (média + erro padrão) para as 4 estações analisadas.
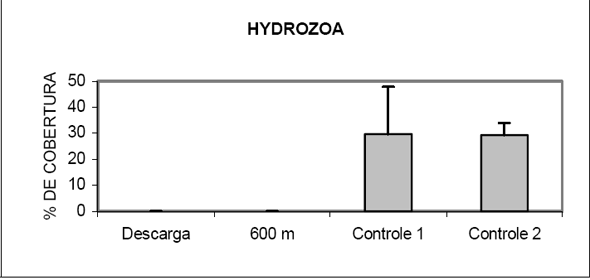
Embora os resultados obtidos tenham abordado uma série temporal única e curta; o que não é indicado para estudos de comunidades, ficou evidente que a descarga do efluente de refrigeração da CNAAA gera impacto no Saco Piraquara de Fora. A região impactada em questão apresentou menor recrutamento e menor riqueza de espécies. Nesta região, a estação mais afastada da saída do efluente apresentou um aumento do número de táxons, indicando uma diminuição dos efeitos nocivos da descarga do efluente. O impacto é maior no verão, considerando as maiores temperaturas registradas neste período. Deste modo o impacto detectado na avaliação realizada foi certamente subestimado.
7.7. Unidades de Conservação (UC) - topo
O litoral sudeste do estado do Rio de Janeiro abriga importantes remanescentes de Mata Atlântica protegidos em três Unidades de Conservação principais: a Área de Proteção Ambiental do Cairuçu, a Reserva Ecológica da Juatinga e o Parque Nacional da Serra da Bocaina, sendo esse último a maior Unidade de Conservação Federal que inclui esse tipo de ecossistema. A importância ambiental da região é salientada por Wege & Long (1995), que consideram-na uma das áreas-chave para a conservação da avifauna no Neotrópico. Atualmente, a despeito de sua importância, a riqueza e a diversidade da avifauna local estão bastante ameaçadas e pouco tem sido feito no sentido de preservá-las.
Entre os principais processos de degradação dos ecossistemas florestais da bacia da Baía da Ilha Grande estão o desmatamento, o corte de madeira de lei, a caça, a retirada de palmito e plantas ornamentais, a extração de terra, os aterros, a expansão urbana e as atividades agropecuárias (MMA, 1997). A partir da análise de fotos aéreas e informações da população local, constatou-se que, nas baixadas e encostas adjacentes, a ampliação de áreas de pastagem, o cultivo de monoculturas e o crescimento urbano têm sido os principais fatores responsáveis pela descaracterização da vegetação nos últimos quarenta anos. A situação dessas terras torna-se ainda mais crítica se for considerado que a maior parte de sua área não está inserida em unidades de conservação, além de estar sujeita a uma das mais fortes pressões de especulação imobiliária do Estado.
Atualmente, a região Baía da Ilha Grande é a segunda microrregião em área de Unidades de Conservação do Estado do Rio de Janeiro (Mendonça et al.1996) e tornou-se uma importante área de preservação a partir da criação das seguintes Unidades de Conservação: Parque Estadual da Ilha Grande (Decreto Estadual 16.067 de 04/06/1973), Reserva Biológica da Praia do Sul (Decreto Estadual 4.972 de 02/12/1981), APAs (Área de Proteção Ambiental) de Cairuçu (Decreto Federal 89.242 de 27/12/1983) e Tamoios (Decreto Estadual 9.452 de 05/12/1986), Reserva Biológica da Ilha Grande (Decreto Estadual 9.728 de 06/03/1987) e Parque Marinho de Aventureiro (Decreto Estadual 15.983 de 27/11/1990).
A região, por possuir alta diversidade e elevado número de espécies endêmicas e/ou ameaçadas de extinção, está incluída em um dos centros brasileiros de diversidade de plantas reconhecidos pela WWF - World Wildlife Fund for Nature e IUCN - The World Conservation Union (Davis et al., 1997). Apesar disso, as espécies desta região, ainda estão pouco representadas nos levantamentos, demonstrando lacunas no conhecimento científico (Marques, 1997).
A caracterização destas Unidades de Conservação da AII-50 km encontra-se no Volume 4 (Diagnóstico Ambiental - Meio Socioeconômico) deste EIA, juntamente com o mapa de localização destas UCs.
|